INTRODUCCIÓN
El Parque Nacional Cabo Pulmo (PNCP) fue establecido en 1995 con el fin de proteger al arrecife de coral más septentrional del Pacífico oriental tropical. Este es un sitio de alta diversidad, donde se han registrado 328 especies de invertebrados y 226 de peces (Villarreal Cavazos et al. 2000; CONANP 2006). La existencia del arrecife se explica porque el sur del Golfo de California recibe reclutas coralinos de diversas regiones del Pacífico mexicano (Chávez Romo et al. 2009; Paz García et al. 2009; Saavedra Sotelo et al. 2011), estos pueden desarrollarse exitosamente, gracias a la presencia de concentraciones relativamente bajas de nutrientes, temperatura adecuada y gran área de plataforma rocosa disponible para el asentamiento larval (Halfar et al. 2004; Reyes Bonilla y López Pérez 2009).
El Área Natural Protegida (ANP) cuenta con 12 especies constructoras de arrecifes, de los géneros Pocillopora, Pavona, Porites, Psammocora y Fungia (Reyes Bonilla y López Pérez 2009). Su cobertura fluctuó de 40% en la década de los 80 (Reyes Bonilla 1993b; Arizpe Covarrubias 2004), a entre el 20% y 30% en la década de los 90 (Reyes Bonilla y Calderón Aguilera 1999); dicha disminución fue causada por el evento de Oscilación Sureña de El Niño de 1997, que causó blanqueamientos y fuertes mortalidades (Reyes Bonilla 2001; Reyes Bonilla y Alvarez Filip 2009). La baja continuó en la década del 2000 debido al impacto de varios huracanes (Alvarez Filip et al. 2006; Calderón Aguilera et al. 2007). Finalmente, Reyes Bonilla y López Pérez (2009) dan el valor del 23% de cobertura a finales de la primera década de este siglo.
Considerando que la protección al arrecife es uno de los objetivos prioritarios del ANP, en este proyecto se realiza una evaluación actualizada de la abundancia coralina y la condición de salud de los corales en el PNCP, además de dar información sobre la estructura de tallas y salud general de este grupo de organismos, y sobre las condiciones ambientales imperantes en la zona. Todo esto será un insumo importante para el manejo del área protegida.
Características ambientales de la zona: El Parque Nacional Cabo Pulmo está ubicado en el Municipio de Los Cabos, Baja California Sur (entre las ciudades de La Paz y San José del Cabo), entre los 23° 22’ 30’’ y los 23° 30’ 00’’ norte y los 109° 28’ 03’’ y los 109° 23’ 00’’ oeste, en la costa del Golfo de California. Su superficie es de 7,111-01-00 ha (Figura 1). El 99% de su extensión está constituida por la porción marina y el 1% es la zona federal marítimo terrestre.
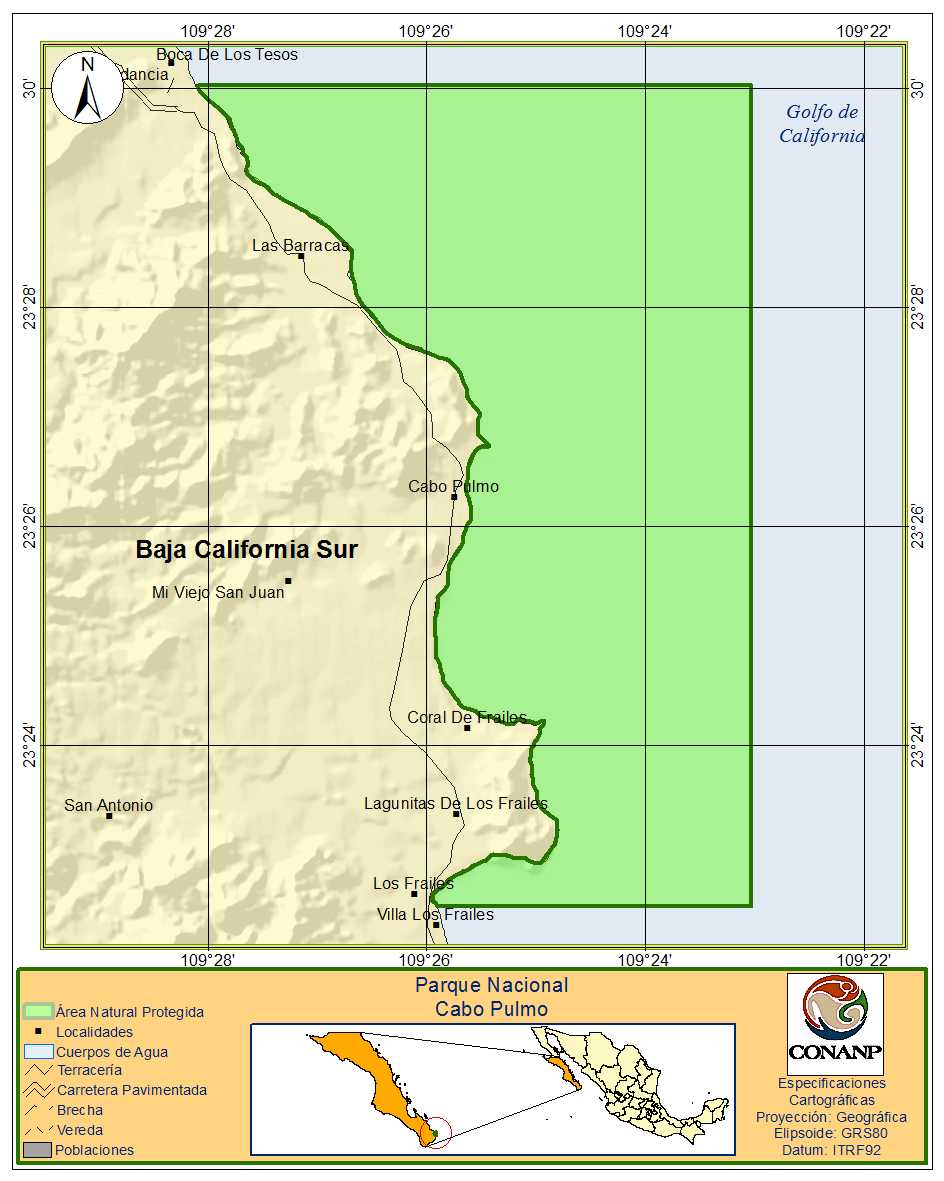 Figura 1. Ubicación del Parque Nacional Cabo Pulmo, en el Golfo de California.
Figura 1. Ubicación del Parque Nacional Cabo Pulmo, en el Golfo de California.
El clima característico de la región es muy seco y cálido, con régimen de lluvias de verano y una precipitación invernal de aproximadamente el 10% del total anual, es importante señalar que la precipitación es muy escasa durante todo el año. El agua dulce en la zona de influencia del Parque y superficies aledañas es escasa y limitada durante los meses de sequía (mayo-octubre), por lo que se aprovecha en forma racional. Este es uno de los factores que limitan la diversificación de las actividades productivas. El área corresponde a la región hidrogeológica número seis y hay en ella dos cuencas la del arroyo Trinidad y la del arroyo San José, que vierten sus aguas en el Golfo de California. Las corrientes que drenan ambas cuencas son intermitentes y con patrones de drenaje integrados (CONANP 2006).
Características biológicas de la zona: Los esfuerzos encaminados a conocer la flora marina del arrecife coralino han sido escasos. Solo se encuentra un estudio (Anaya y Riosmena 1996). La extracción de especímenes se realiza prácticamente con fines científicos. Aún cuando se presentan especies de valor comercial, no existen reportes de aprovechamiento económico. Entre las especies de importancia comercial que se han registrado en el Parque Nacional, están Sargassum horridum, Sargassum sinicola, Gracilaria spinigera, Ulva lactuca, Halymenia californica, Halymenia templetonii, Hypnea johnstonii y Hypnea cervicornis. Otras especies de algas tienen importancia ecológica, ya que dentro de la comunidad ficológica constituyen mantos de muy amplia cobertura, en donde ocurre una red de complejas interacciones sucesionales entre estas plantas y los organismos formadores del arrecife. En el caso de la vegetación terrestre, la comunidad predominante está constituida por matorral sarcocaule con las siguientes especies: Jatropha cinerea, J. cuneata, Fouquieria sp. y Machaerocereus gummosus (CONANP 2006). De su fauna, el arrecife tiene una edad aproximada de 20,000 años, antigüedad que comparada con otros arrecifes de América, la ubica dentro de los más viejos del Pacífico americano ya que los de Panamá, por ejemplo, tienen apenas 5,000-5,500 años (Glynn y McIntyre 1977). A pesar de que, en general, la fauna arrecifal está mejor estudiada que su flora, el conocimiento de ella es aún insuficiente. Actualmente, los peces, corales y moluscos están mejor estudiados que el resto de los animales. El arrecife del Parque presenta la cobertura coralina más extensa en el Golfo de California, y en él habitan 11 de las 14 especies de corales hermatípicos reportados para el golfo: Pocillopora verrucosa, Pocillopora capitata, Pocillopora damicornis, Pocillopora meandrina, Pavona gigantea, Pavona clivosa, Porites panamensis, Psammocora stellata, Psammocora brighami, Fungia curvata, Madracis pharensis (Reyes Bonilla 1993a), todos estos son considerados como corales duros. De igual forma, en el caso de la comunidad ictiológica se tienen observaciones de 226 especies arrecifales (Villarreal 1988) de las 875 especies listadas para el Golfo de California (Finley et al. 1996). Otro grupo ampliamente representado en el arrecife es el de los moluscos, algunas de las especies identificadas son: Conus brunneus, Conus princeps, Murex elenensis, Muricanthus princeps, Thais kiosquiformis y Pinctada mazatlanica. Estos organismos tienen importancia comercial, algunos como piezas de colección llegan a cotizarse en muy altos precios y otras especies son valiosas por los subproductos que se obtienen de ellas, además de estar presentes en la lista de especies en peligro de extinción, como es el caso de la madre perla (Pinctada mazatlanica). Otros moluscos tienen gran demanda por su excelente sabor. En el área están presentes especies bajo alguna categoría de protección por las leyes mexicanas (NOM-059-SEMARNAT- 2010), como las tortugas que visitan algunas playas: Dermochelys coriacea, C. caretta, Lepidochelys olivacea, Chelonia agassizi y Eretmochelys imbricata. Las aves marinas presentes en el Parque son comunes en todo el golfo, incluye a Larus livens, L. hermanni, Sterna antillarum, S. elegans, S. maxima, Pelecanus occidentalis, Ardea herodias, A. alba, Limosa fedoa, Numenius phaeopus y N. americanus. En el Parque se observa una colonia de Zalophus californianus. Cerca de la costa se pueden observar otros mamíferos marinos que transitan dentro y cerca de los límites del Parque, como Tursiops truncatus, Stenella longirostris y Steno bredanensis. En invierno se observan a Megaptera novaeangliae, Balaenoptera physalus y B. edeni (Figura 2; CONANP 2006).
El objetivo principal fue elaborar un diagnóstico de la cobertura actual de las especies de coral en el Parque Nacional Cabo Pulmo, así como determinar la cobertura coralina de las especies del género Pocillopora, detectar la presencia y extensión del blanqueamiento coralino, enfermedades o daños por actividades antrópicas, determinar el estado de conservación general de la estructura coralina, identificar las zonas coralinas mejor conservadas y las que son susceptibles de restauración y proporcionar a las autoridades del parque, información oceanográfica actualizada de la zona de estudio, tomada de imágenes satelitales.
 Figura 2. Corales y peces en el Parque Nacional Cabo Pulmo. Figura 2. Corales y peces en el Parque Nacional Cabo Pulmo.
MONITOREO
Metodología: Los resultados que se presentan se obtuvieron con base en trabajo de campo y consulta de información de bases de datos oceanográficas internacionales. En el primer caso, el PNCP fue visitado en dos ocasiones; la primera fue del 16 al 19 de agosto, y la segunda del 13 al 16 de noviembre de 2011, fechas en que se recorrieron las 11 áreas indicadas en el mapa (Figura 3). En cada una se hicieron entre 5 y 6 transectos de banda de 20 m de largo por 1 de ancho (N=122), dentro de los cuales se estimó el porcentaje de cobertura coralina por especie dentro de cuadrantes de 1 m2, divididos en 100 secciones de 100 cm2 cada una (English et al. 1997; Raymundo et al. 2008).
Una vez con la información sobre la cobertura coralina por especie, para cada transecto se calcularon los valores de los siguientes índices ecológicos: riqueza (número de especies), abundancia (o cobertura), el índice de diversidad de Shannon-Wiener (H´, en base 10) y el índice de uniformidad de Pielou (J´). Las cifras de cada índice fueron comparadas entre sitios usando análisis de varianza de una vía, modelo II, y en el caso de encontrarse diferencias significativas, se usó la prueba de Tukey para detectar el origen de las diferencias estadísticas.
Por otra parte, para diagnosticar la cobertura viva de corales en todo el parque, se emplearon datos generados en un proyecto sobre mapeo de los principales hábitat del arrecife donde colabora el responsable de la presente propuesta. Para obtener la información, se llevaron a cabo arrastres manta (“manta tow”) en dirección perpendicular a la playa y a una velocidad de 3 a 5 nudos. El buceador tomó información sobre la cobertura de coral cada 30 segundos, anotando datos en tablillas de acrílico, y basándose en un área de observación circular de aproximadamente 5 m de diámetro (Hill y Wilkinson 2004). Con este procedimiento, la totalidad del parque (profundidades de 0 a -30 m) fue mapeada a una escala de 1:20,000, y posteriormente con un sistema de información geográfica se estimó la extensión total en hectáreas que cubre el coral vivo. Además, se estimó el tamaño de los 11 arrecifes designados en la convocatoria como “Zonas de importancia” (Figura 3).
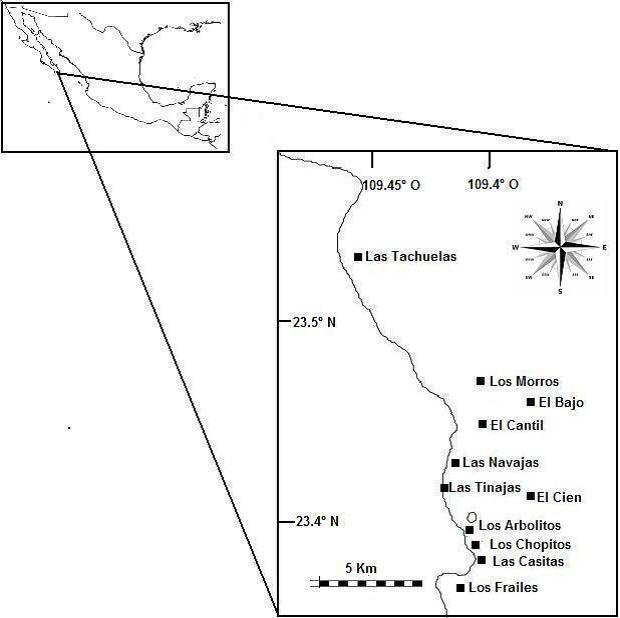
Figura 3. Área de estudio, mostrando los sitios visitados en el PNCP.
El tercer análisis correspondió a la revisión de la estructura de tallas de las especies presentes, para ello durante las salidas de campo se determinó la altura y el diámetro mayor (precisión + 1 cm) de 523 colonias coralinas (154 en agosto y 369 en noviembre) que estaban en posición de crecimiento original, o bien, fragmentos cementados sobre rocas o anclados sobre bases de esqueleto carbonatado. Cada colonia fue revisada para denotar la presencia de blanqueamiento, enfermedades y daños antropogénicos, y se cuantificó el porcentaje de tejido muerto, visto desde arriba.
Para definir el estado actual de conservación de los 11 sitios visitados, se hizo un análisis multicriterio. Se tomaron en cuenta los valores de los cuatro índices ecológicos calculados (S, N, H´, J´) y de los indicadores de perturbación, y estos fueron ranqueados asignando el valor de 11 al lugar en mejor condición (más alta riqueza, cobertura, diversidad y uniformidad; o menor porcentaje de mortalidad, blanqueo, enfermedades y fragmentación), y de 1 al que tuvo el peor estado. Al final, se sumaron los rangos y con base en ellos se denotaron los valores de calidad relativa para cada sitio.
Finalmente, de la base de datos Giovanni (Ocean Color Radiometry Online Visualization and Analysis), administrada por la National Aeronautic and Space Agency (NASA), se obtuvo la serie de tiempo del valor del coeficiente de atenuación de la luz y clorofila (enero de 1998 a diciembre de 2010) con base en datos del satélite SEAWIFS, así como de la temperatura superficial del agua (enero 2002 a diciembre 2010), tomados del satélite MODIS-AQUA. La información se presenta en unidades de 0.01° de latitud-longitud (aproximadamente 100 m de lado), y en forma de mapas.
Resultados: Para la determinación de la cobertura coralina se incluye la información de su abundancia junto con la de la estructura comunitaria de este grupo en el parque. Los datos son relevantes en sí mismos y además son fundamentales en la evaluación de la condición actual de los distintos sitios en la zona.
La cobertura total estimada en los 122 transectos fue del 12.46 + 1.10% del fondo (promedio y error típico; Figura 4), siendo mucho más alta en Las Tachuelas (37.65 + 4.22 %), seguida de El Cantil (19.30 + 2.86 %), mientras que de manera opuesta en El Cien la cifra fue la más baja (1.31 + 0.23 %). El análisis de varianza demostró que hay diferencias significativas entre sitios (F10,111= 18.96, p< 0.01) y la prueba a posteriori mostró tres grupos respecto a los niveles de cobertura: Las Tachuelas (>20%), seguido por El Cantil-Las Navajas-El Bajo-Las Tinajitas-Las Casitas (10%-20%), y al final el resto (<10%).
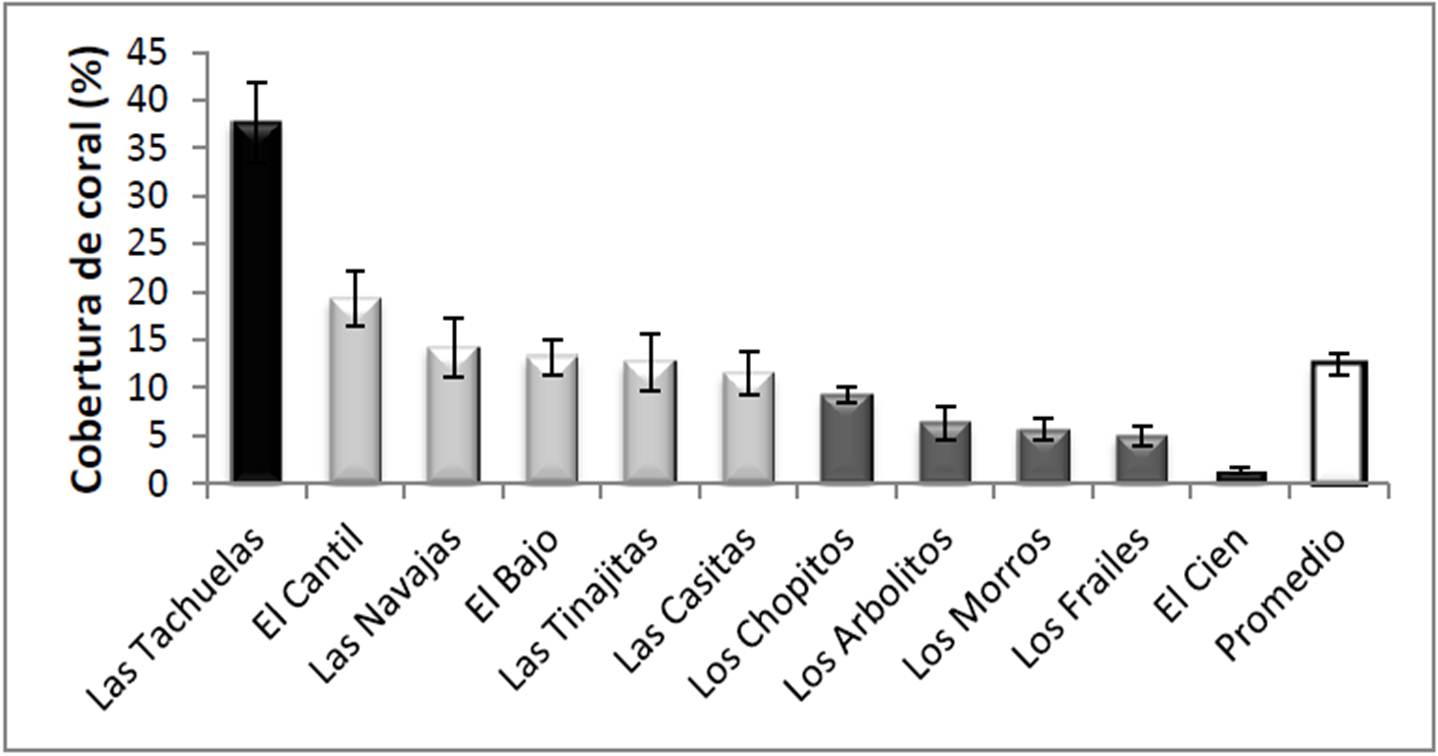
Figura 4. Cobertura de corales en los sitios visitados en el PNCP, y promedio general de todo el parque.
Los valores de cobertura en el arrecife referidos en la literatura para años recientes superan el 20%, sin embargo esto se debe a que los análisis se realizaron solo en áreas someras (menos de 12 m). Esto es relevante dado que en Cabo Pulmo la cobertura depende en gran medida de la profundidad del sitio (Figura 5), ya que el coral requiere mucha luz para el adecuado desarrollo de sus zooxantelas (Iglesias Prieto et al. 2004). Así, si se excluyen los lugares con profundidad de 10 m o más (El Bajo, Los Morros y El Cien), la cobertura coralina del PNCP pasa a ser del 14.56 + 1.40%, significativamente menor que un 20%. La cifra estimada en agosto para las zonas someras fue del 16.93 + 3.14%, es decir, un 2.37% más alta que la medida en noviembre. Esta diferencia, como se verá adelante, es resultado de la ocurrencia en otoño de uno de los tres principales eventos de blanqueamiento coralino registrados históricamente en la zona.
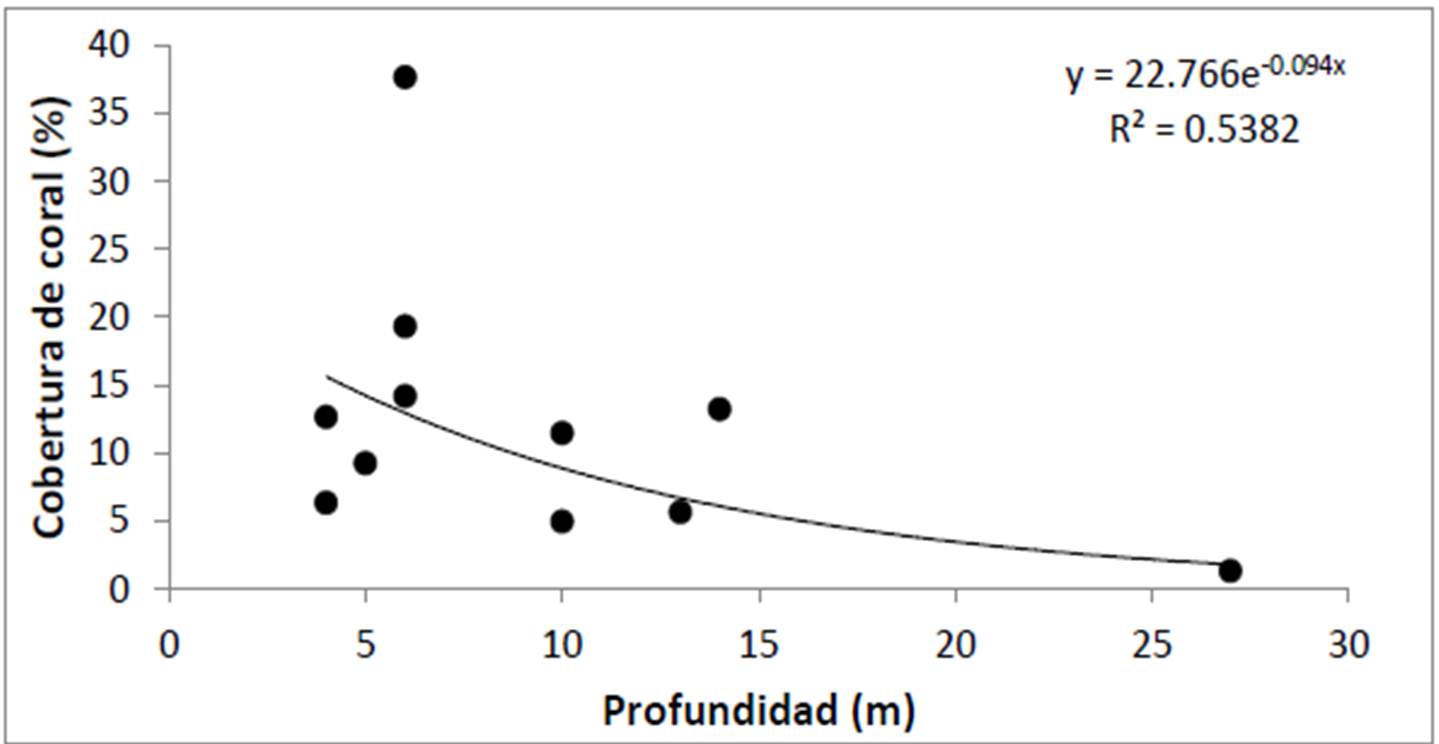 Figura 5. Relación entre la profundidad y la abundancia coralina en el PNCP, tomando en cuenta el promedio de cobertura por sitio.
Figura 5. Relación entre la profundidad y la abundancia coralina en el PNCP, tomando en cuenta el promedio de cobertura por sitio.
En relación con la abundancia relativa por especie, los corales Pocillopora meandrina y P. verrucosa fueron dominantes con un 34% y 33% respectivamente de la cantidad total de cobertura viva (Figura 6). Le siguieron Porites panamensis y Pavona gigantea (entre 11% y 9% de la cobertura coralina relativa) y en conjunto estas especies suman más del 85% de toda la cobertura coralina sobre el fondo en el parque. La tendencia a que las cuatro especies citadas sean las principales constructoras del arrecife se ha mantenido desde la década de los 90 (Reyes-Bonilla 1993b), indicando su resiliencia a repetidas perturbaciones como huracanes, eventos de El Niño y blanqueamientos coralinos.
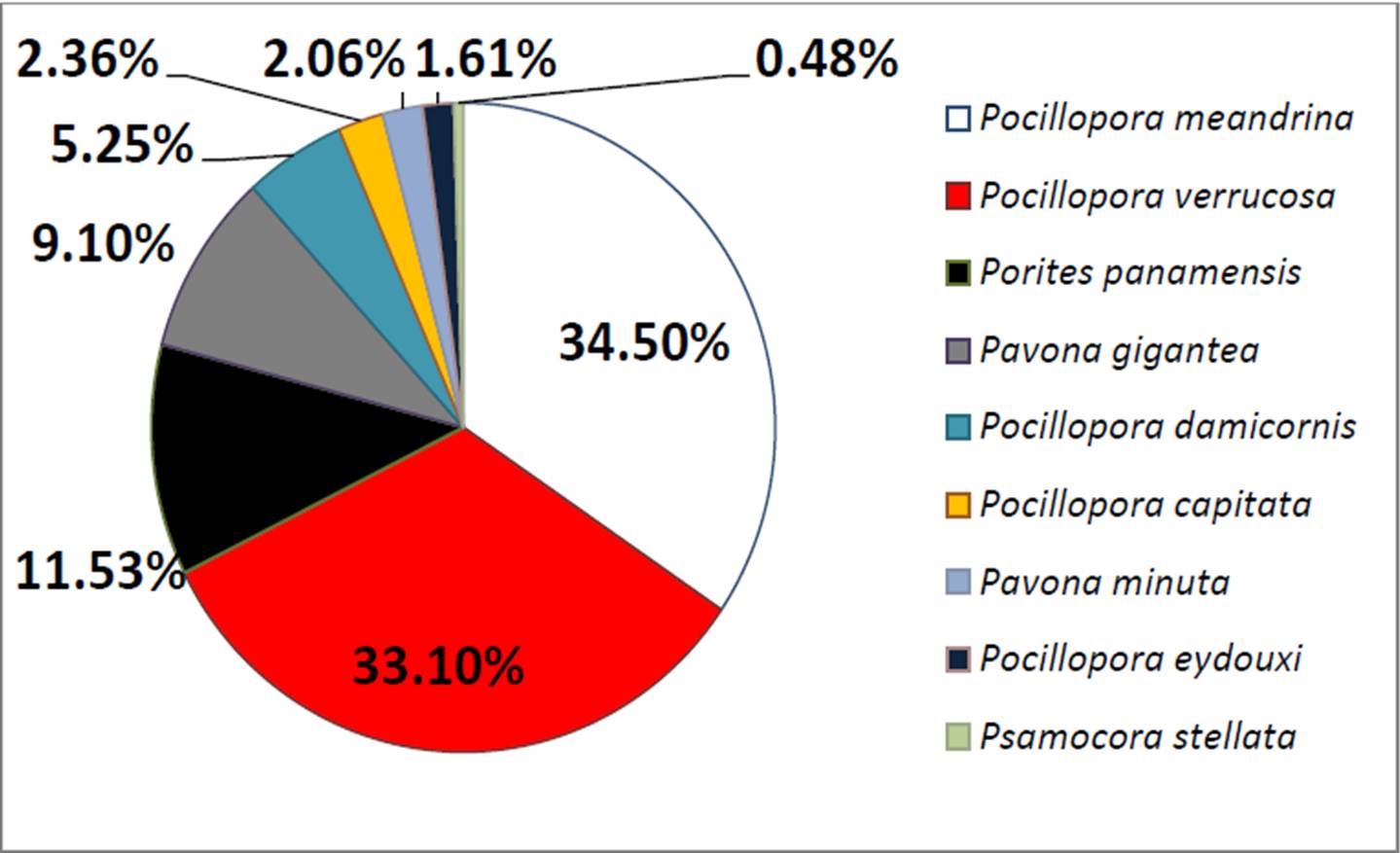 Figura 6. Abundancia relativa de las especies de corales zooxantelados del PNCP. La cobertura total que suman entre ellas es del 14.56% del fondo.
Figura 6. Abundancia relativa de las especies de corales zooxantelados del PNCP. La cobertura total que suman entre ellas es del 14.56% del fondo.
En lo que se refiere a la riqueza coralina, en el PNCP se encontraron 9 especies constructoras de arrecife pertenecientes a 4 géneros. En promedio en todo el parque se localizaron 3.09 ± 0.09 spp/censo. Los análisis estadísticos mostraron diferencias significativas entre los sitios (F10,111=8.69, p<0.0001) formando tres grupos, El Cien como el de menor riqueza con menos de 2 spp/censo (1.2 ± 0.13 spp/censo), seguido del grupo formado por Los Frailes-Los Morros-Las Casitas con ≤3 spp/censo, y finalmente el resto de los sitios con más de 3 spp/censo, siendo Los Chopitos el de mayor riqueza (4.16 ± 0.29 spp/censo; Figura 7). En resumen, puede decirse que la riqueza por transecto es muy homogénea en el PNCP, con valores muy altos en ciertas áreas por ser someras (Los Chopitos) o estar conservadas (El Bajo, El Cantil), o muy bajos en sitios especiales dada su gran profundidad (El Cien).
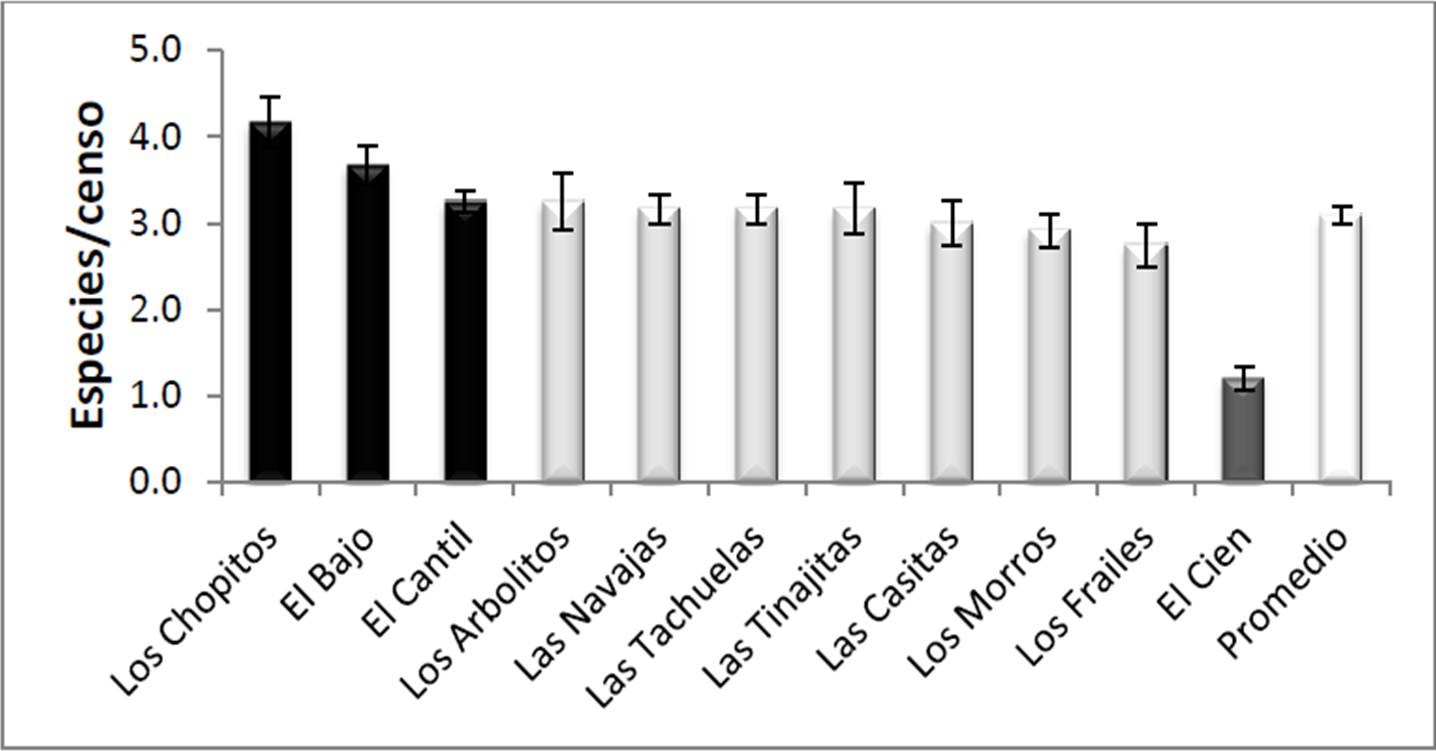 Figura 7. Riqueza de corales (promedio ± error típico) en cada uno de los sitios visitados en el PNCP, y promedio general de todo el parque. Con distintos tonos de gris se señalan los grupos definidos por el análisis de varianza y las pruebas a posteriori.
Figura 7. Riqueza de corales (promedio ± error típico) en cada uno de los sitios visitados en el PNCP, y promedio general de todo el parque. Con distintos tonos de gris se señalan los grupos definidos por el análisis de varianza y las pruebas a posteriori.
En cuanto a diversidad se refiere (H´), el PNCP mostró una diversidad promedio de 0.34 ± 0.01, siendo Los Chopitos el sitio más diverso de todos (0.48 ± 0.03), mientras que de manera contraria, El Cien fue el menos diverso (0.05 ± 0.03). Los análisis estadísticos indicaron diferencias significativas entre los sitios (F10,111=7.355, p<0.0001), formando claramente tres grupos; Los Chopitos y El Bajo como los más diversos, El Cien como el menos diverso y finalmente el resto de los sitios con un valor intermedio (Figura 8).
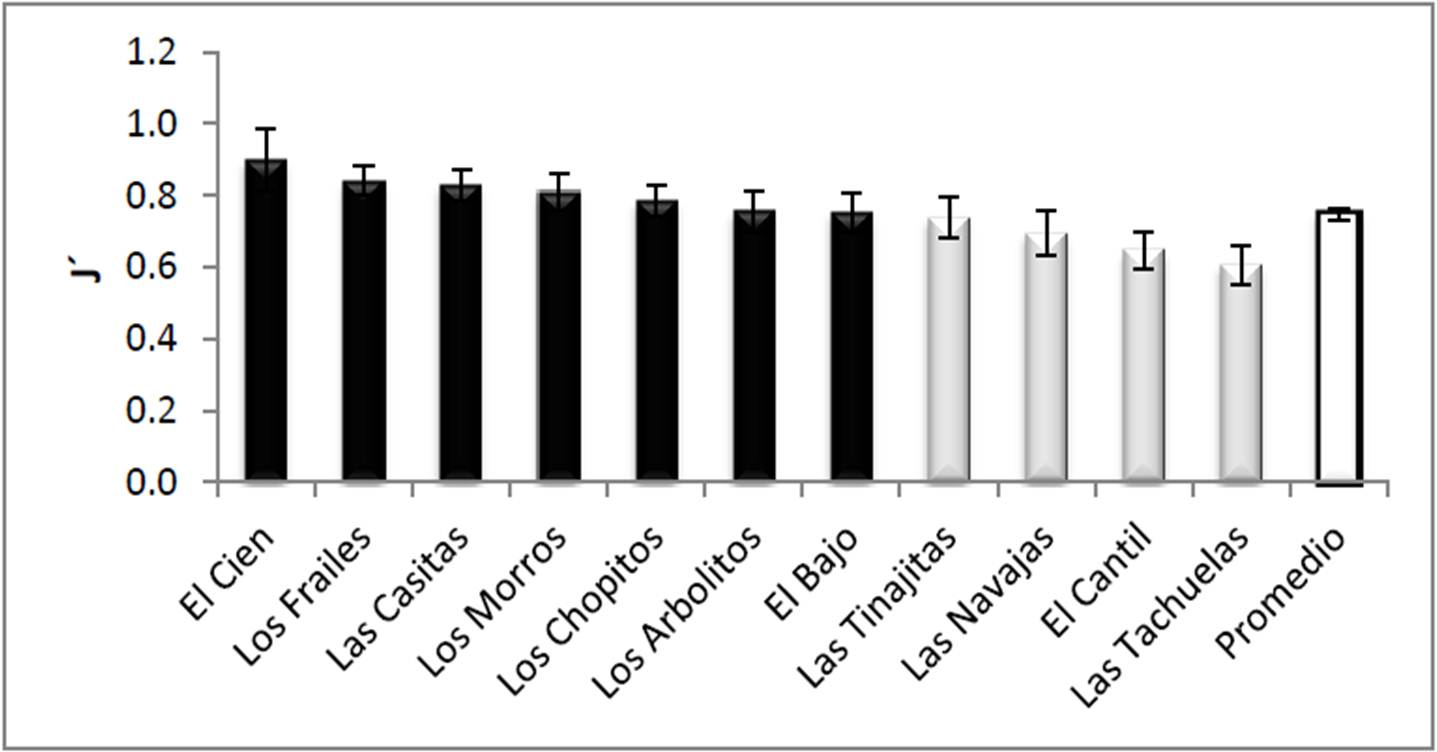 Figura 8. Equidad de corales (promedio ± error típico) en los sitios visitados del PNCP, y promedio general de todo el parque. Con distintos tonos de gris se señalan los grupos definidos por el análisis de varianza y las pruebas a posteriori.
Figura 8. Equidad de corales (promedio ± error típico) en los sitios visitados del PNCP, y promedio general de todo el parque. Con distintos tonos de gris se señalan los grupos definidos por el análisis de varianza y las pruebas a posteriori.
La equidad es un indicador de la proporción relativa de la abundancia de las especies presentes en un sitio, y toma valores más altos (cercanos a 1) si esta es similar o si no existe dominancia marcada de un taxón en especial. Al contrario, se acerca a cero cuando uno o pocos grupos tienen abundancias desproporcionadas en relación con los demás.
La equidad de corales del PNCP fue de 0.74 ± 0.01 en promedio (Figura 9), donde el sitio conocido como El Cien fue el más uniforme de todos (0.89 ± 0.08); por otro lado el área con menor uniformidad del parque fue Las Tachuelas (0.60 ± 0.05). Estos valores, junto con los datos de la Figura 6, muestran que la comunidad coralina en el parque mantiene un cierto equilibrio en relación con la cobertura de las especies constituyentes. Finalmente, los estadísticos diferenciaron dos grupos (F10,11=2.24, p=0.021), Las Tachuelas, El Cantil, Las Navajas y Las Tinajitas (todos ellos situados al norte del PNCP) como los sitios menos uniformes, y el resto (al sur) tuvieron la mayor equidad (Figura 9).
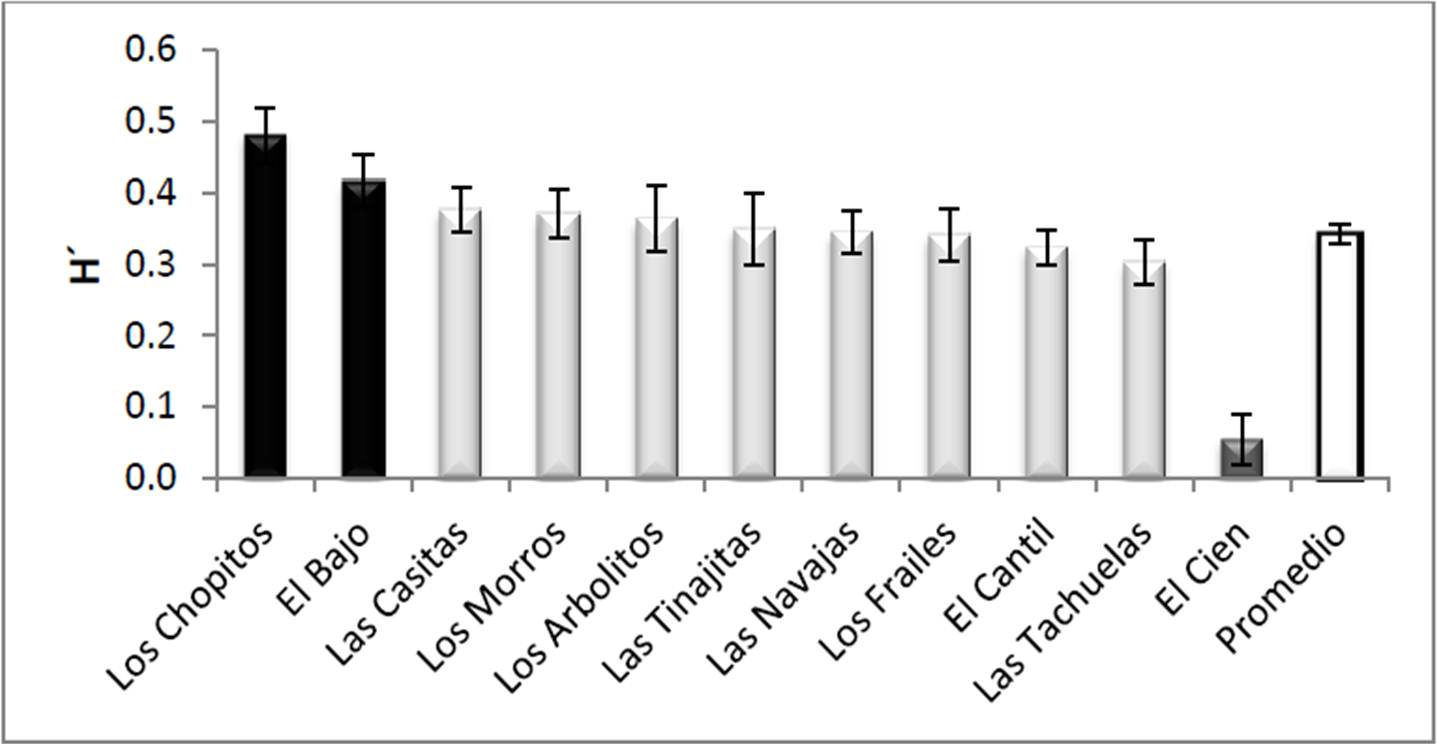
Figura 9. Diversidad de corales (promedio ± error típico) en los sitios visitados del PNCP, y promedio general de todo el parque. Con distintos tonos de gris se señalan los grupos definidos por el análisis de varianza y las pruebas a posteriori.
El mapa de hábitat de fondo de Cabo Pulmo (Figura 10) muestra que la cobertura coralina viva aparece especialmente en el norte. También se pueden detectar los efectos de las mortalidades causadas por ciclones y eventos ENSO cerca de los límites de la bahía en forma de grandes áreas de coral muerto. Por otra parte, la gráfica de pastel (Figura 11) indica que la zona de coral vivo estimada en 2010 por los arrastres de manta equivalía al 21% del área revisada por buceo (323.4 hectáreas de 1,521 en total; las restantes estuvieron a más de 30 m de profundidad y no fueron evaluadas). En resumen, la estimación de 2010 converge alrededor de la cifra del 20% de cobertura coralina indicada en la literatura. También, es interesante anotar que si se suma la superficie de coral muerto y vivo, se tendría un 32% del fondo, que es aproximadamente la cifra dada en los 90 (31%; Reyes Bonilla y Calderón Aguilera, 1999). Desafortunadamente, el valor de cobertura viva sensiblemente disminuye en el otoño de 2011, como se describirá en la siguiente sección.
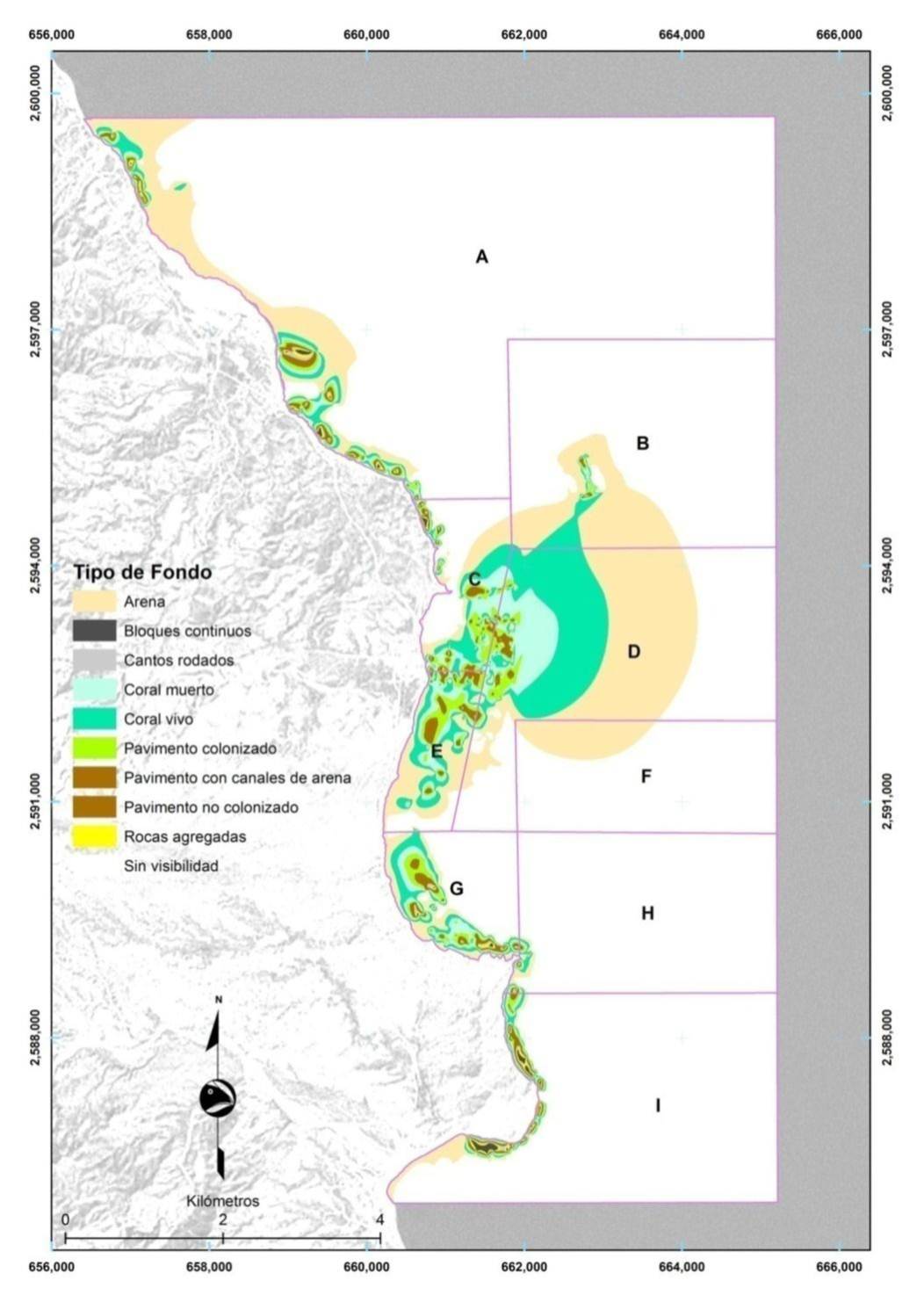 Figura 10. Mapa de tipos de fondo en el PNCP. Las zonas en blanco corresponden a sitios de más de 30 m de profundidad.
Figura 10. Mapa de tipos de fondo en el PNCP. Las zonas en blanco corresponden a sitios de más de 30 m de profundidad.
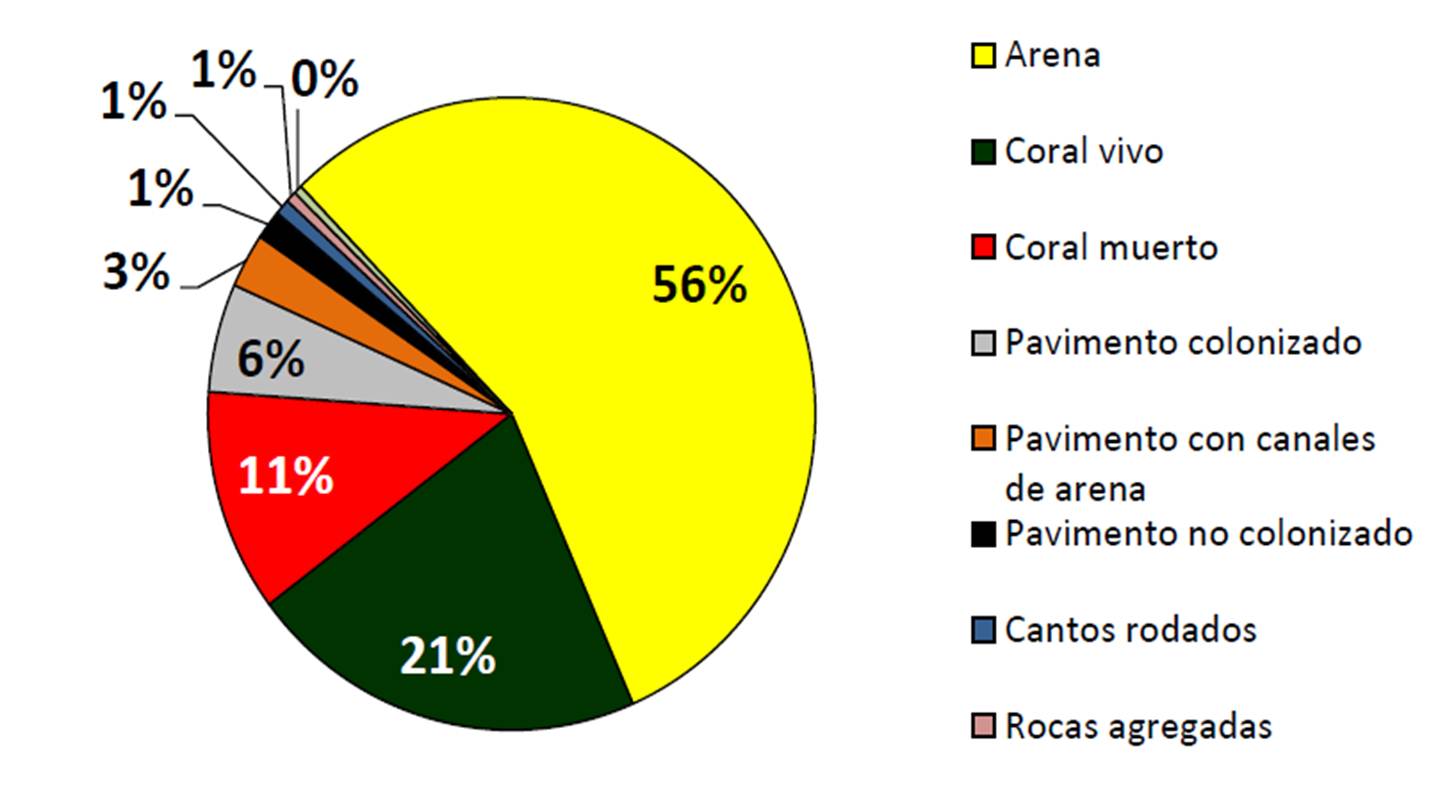 Figura 11. Área relativa del PNCP ocupada por diversos tipos de fondo (datos de 2010).
Figura 11. Área relativa del PNCP ocupada por diversos tipos de fondo (datos de 2010).
Para detectar la presencia y extensión de enfermedades de corales, daños por actividades antrópicas o fenómenos de blanqueamiento coralino en el PNCP, se revisaron 523 colonias de 11 especies de coral (154 en agosto y 369 en noviembre), en los 11 sitios visitados en el PNCP. Los resultados muestran una situación muy aceptable dado que no se detectaron enfermedades en el parque, y ello puede explicarse si se considera que estos problemas son extremadamente raros en el Pacífico oriental tropical, habiendo solo dos ejemplos conocidos: la enfermedad de banda blanca y plaga blanca en Colombia (Navas Camacho et al. 2010), y la trematodiasis en colonias de Porites spp., en Manzanillo, México. Es probable que las relativamente bajas temperaturas imperantes en el Golfo de California, comparadas con las que se presentan en el Mar Caribe, el Pacífico tropical o el Indo Pacífico, sean los agentes que evitan la propagación de patógenos.
En relación con la fragmentación aparentemente causada por acción humana (Figura 12), se encontraron 63 colonias afectadas por este problema (12.04% del total), el cual fue particularmente severo en El Bajo (25.9% de las colonias tenían evidencia de ruptura), Los Morros (25.5%) y las Tachuelas (18.4%). Estas tres zonas son frecuentemente visitadas por buceadores libres o con equipo autónomo, lo cual puede explicar el patrón. Al mismo tiempo, los números llaman la atención sobre que ya puede detectarse un efecto de la presencia turística de los sitios sobre las poblaciones de corales, aun cuando el nivel de uso es todavía limitado.
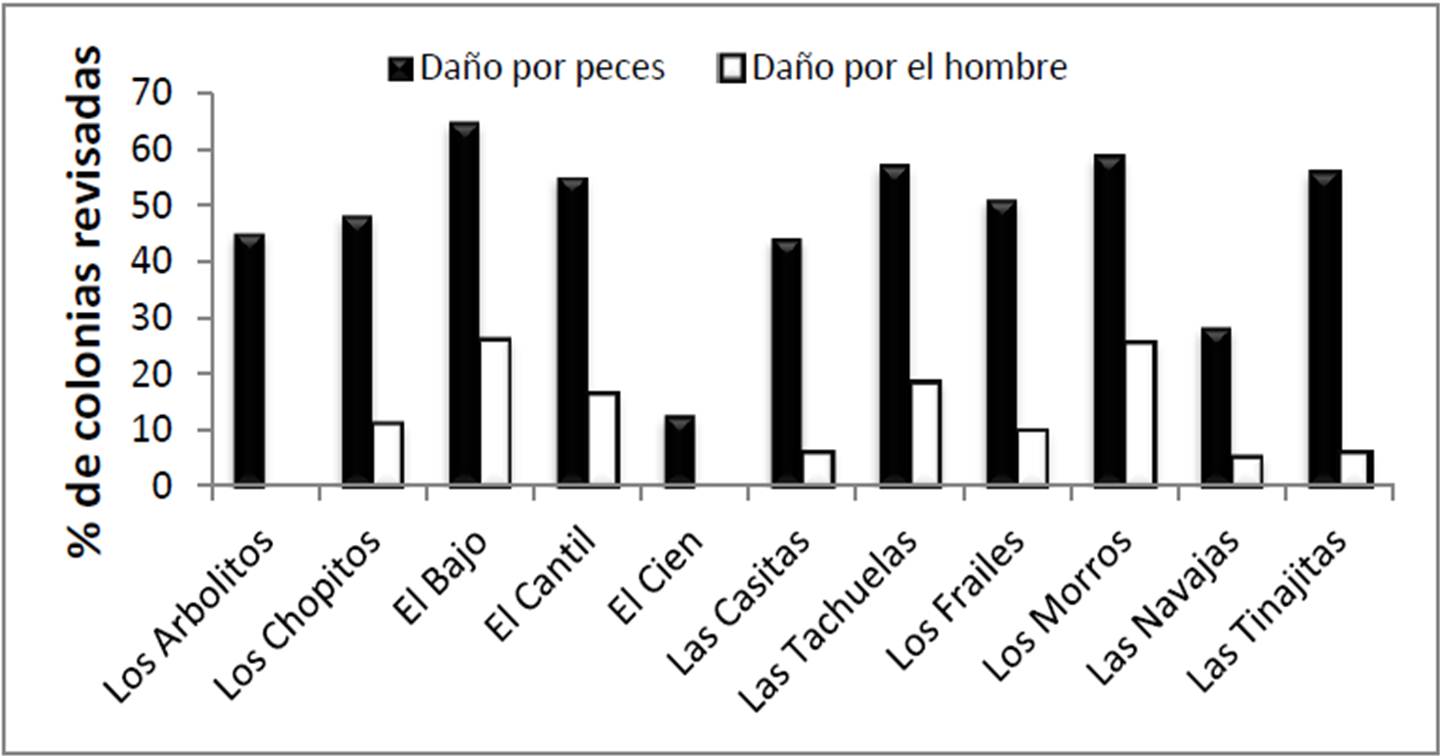
Figura 12. Intensidad relativa de daño por depredación de peces y fragmentación inducida por el hombre en el PNCP.
Por otro lado, el daño causado a los corales por los peces fue ubicuo; el 50.1% de las colonias revisadas (262 de 523; Figura 12) presentó evidencia de mordidas en la parte superior de las ramas (Pocillopora spp., Psammocora spp.) o bien en toda la extensión del corallum (Pavona spp., Porites panamensis). Las zonas principalmente afectadas fueron El Bajo (64.8%), Los Morros (58.8%), Las Tachuelas (57.1%), Las Tinajitas (56.0%), El Cantil (55.1%) y Los Frailes (50.9%), mientras que El Cien presentó la cifra más modesta (12.5%); en resumen, puede verse una tendencia a que el consumo coralino por peces ocurre en la zona norte del parque. Según Reyes Bonilla y Calderón Aguilera (1999), y Moreno y colaboradores (2009), en la zona de Cabo Pulmo los depredadores coralinos por excelencia pertenecen a las familias Scaridae (pericos), Balistidae (cochitos) y Tetraodontidae (botetes), taxa que en efecto fueron abundantes a todo lo largo del parque. No obstante su frecuencia de ocurrencia del daño por depredación no debe representar preocupación dado que es un evento natural y además, el carbonato excretado por los peces en forma pulverizada se transforma en material que ayuda a cementar la estructura arrecifal (Sheppard et al. 2009).
Debido a la relevancia del blanqueamiento coralino de 2011, se decidió abrir un apartado específico para este tema, ya que el evento ocurrido en 2011 es uno de los tres más graves que se han registrado en el arrecife de Cabo Pulmo desde 1987 y quizá, uno de los principales en los últimos 50 años.
Durante la salida de campo de agosto, el blanqueamiento coralino fue nulo, aunque hubo decoloración parcial (calificada como tal cuando el coral mantiene su color verde o café, pero en una tonalidad muy tenue; Figura 13): 38 colonias revisadas (25% del total) sufrieron ese problema, sobre todo en las zonas de Los Chopitos y Las Casitas, que están entre las más someras visitadas. En el Golfo de California la decoloración parcial de los corales no es rara y ocurre durante verano en colonias residentes de agua somera, ya que las zooxantelas inhiben su crecimiento dada la alta irradiación y la elevada temperatura (Iglesias Prieto et al. 2004). En específico en Cabo Pulmo, cada año durante septiembre suelen presentarse ambas situaciones (ver sección de Datos Oceanográficos), y los corales pierden un tanto su color. No obstante, en agosto de 2011 no se presentó verdadero blanqueamiento en el PNCP (pérdida total de simbiontes) porque la temperatura medida por satélite en el sitio durante enero a julio de 2011 no superó los 30.3°C, que es el umbral de blanqueamiento en la zona.
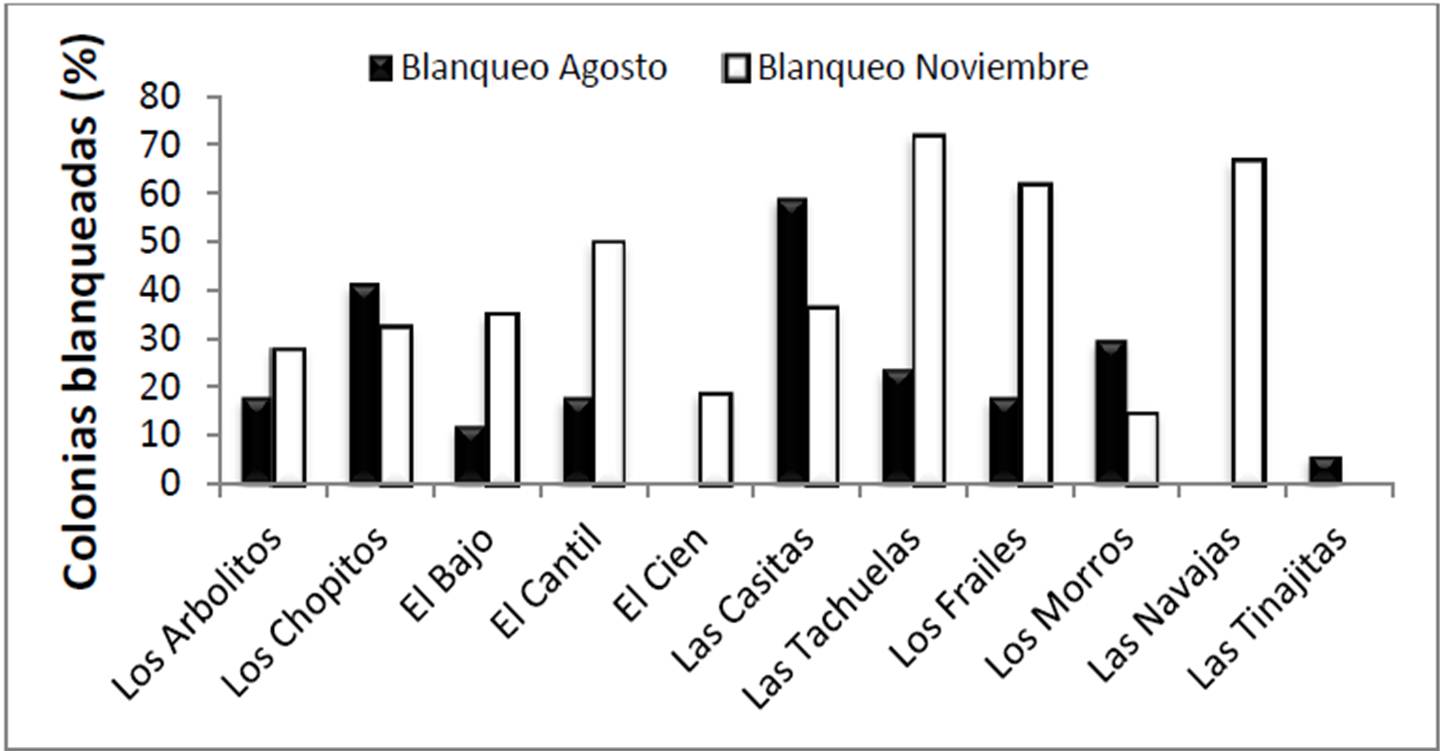
Figura 13. Intensidad relativa de blanqueamiento coralino en agosto (parcial) y noviembre (total) en 2011 en el PNCP.
En contraste con lo anterior, la visita de noviembre mostró 153 colonias (39% de total) con blanqueamiento total, es decir un aumento del 14% de corales afectados respecto a agosto. Se observaron mayores daños en Las Tachuelas (71.9%), Las Navajas (66.7%) y Los Frailes (61.8%), lo que remarca que el daño fue mayor en agua somera que en agua profunda. No obstante ello, incluso en áreas de más de 10 m de profundidad como El Cien, El Cantil y El Bajo se observaron colonias coralinas afectadas. El fenómeno ocurrió principalmente en corales del género Pocillopora (muy posiblemente debido a que reside fundamentalmente en agua somera y que tiende a alcanzar mayor temperatura), pero en realidad todos los géneros sufrieron pérdida de coloración.
En la mayoría de sitios el porcentaje de blanqueo fue mayor en noviembre que en agosto excepto en Los Chopitos, Las Casitas, Las Tinajitas y Los Morros. La disminución del número de colonias afectadas por blanqueamiento parcial y luego total en un mismo sitio ha sido observada antes en la zona de Cabo Pulmo (Reyes Bonilla 1993b; Reyes Bonilla et al. 2002), y no representa una mejora sino que en realidad es lo opuesto: el cambio está relacionado con la mortalidad coralina. Esto se debe a que los corales que pierden primero su coloración tienden a morir en menos de 2 meses debido a que la falta de zooxantelas y la baja cantidad de plancton disponible en aguas someras del sur del Golfo de California en verano, llevan al coral a la inanición y como resultado, el número de corales vivos pero con mengua o pérdida total de color, disminuye en el tiempo. La gráfica del porcentaje de mortalidad coralina parece apoyar esta observación dado que Las Tinajitas y Los Chopitos se encuentran entre los sitios con mayor intensidad de mortalidad local (Figura 14). Sin embargo, esta afirmación debe tomarse con cuidado dado que existe una tasa natural de pérdida de colonias, la cual es desconocida tanto en cantidad total como por sitio.
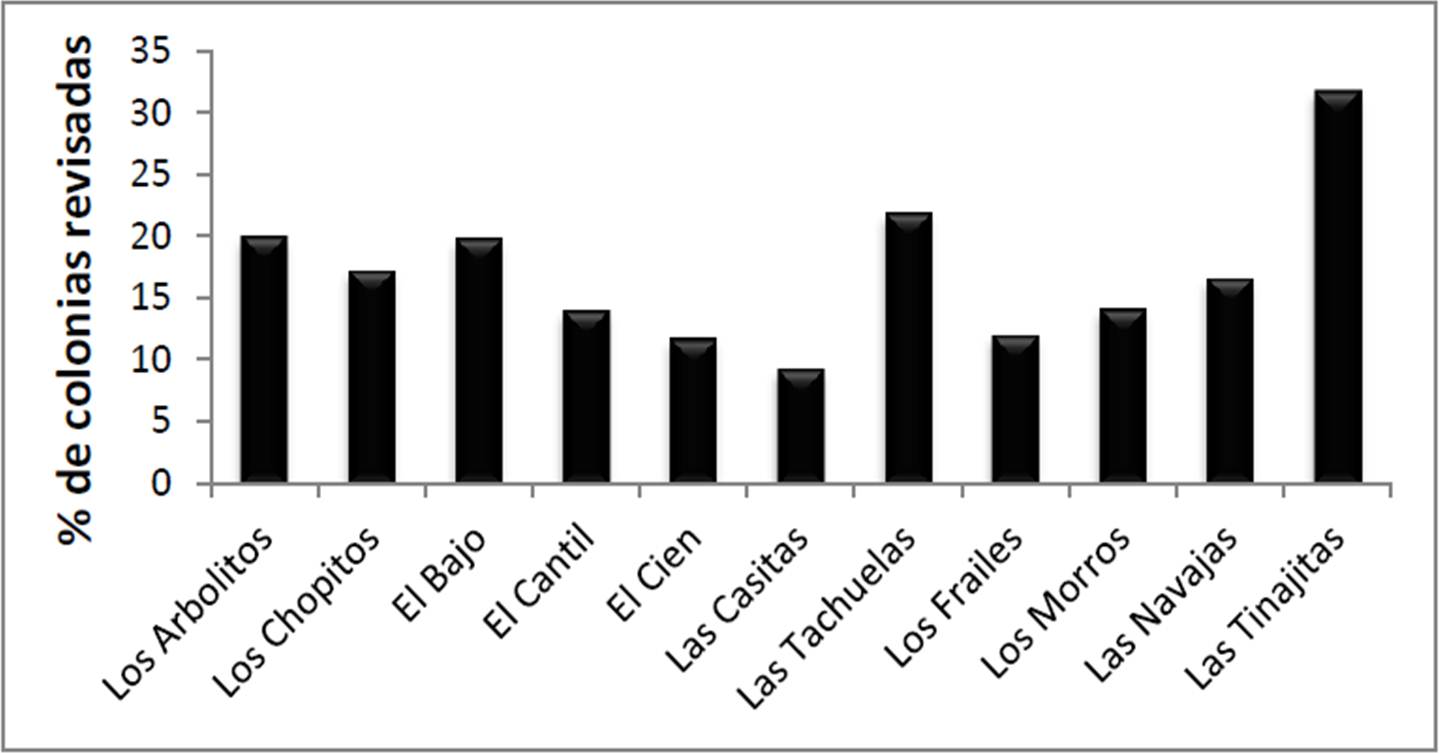 Figura 14. Mortalidad relativa de colonias coralinas en el PNCP.
Figura 14. Mortalidad relativa de colonias coralinas en el PNCP.
Como se indicó en secciones anteriores, la cobertura coralina viva total decreció 2.4% en tres meses a nivel de todo el parque (Figura 4), al analizar por sitios puede verse que hubo tres zonas impactadas por el fenómeno de pérdida de cobertura (muy probablemente relacionado con el blanqueo): Las Tinajitas (-7.62%), El Cantil (-6.76%) y Los Chopitos (-3.05%), todas presentando una disminución significativa de su cobertura coralina en relación con agosto (Figura 15). También puede observarse que prácticamente en todos los sitios la abundancia de coral bajó, excepto en El Bajo (que permaneció casi idéntica) y en Las Tachuelas (que aunque nominalmente aumentó, en realidad la diferencia no fue estadísticamente significativa).
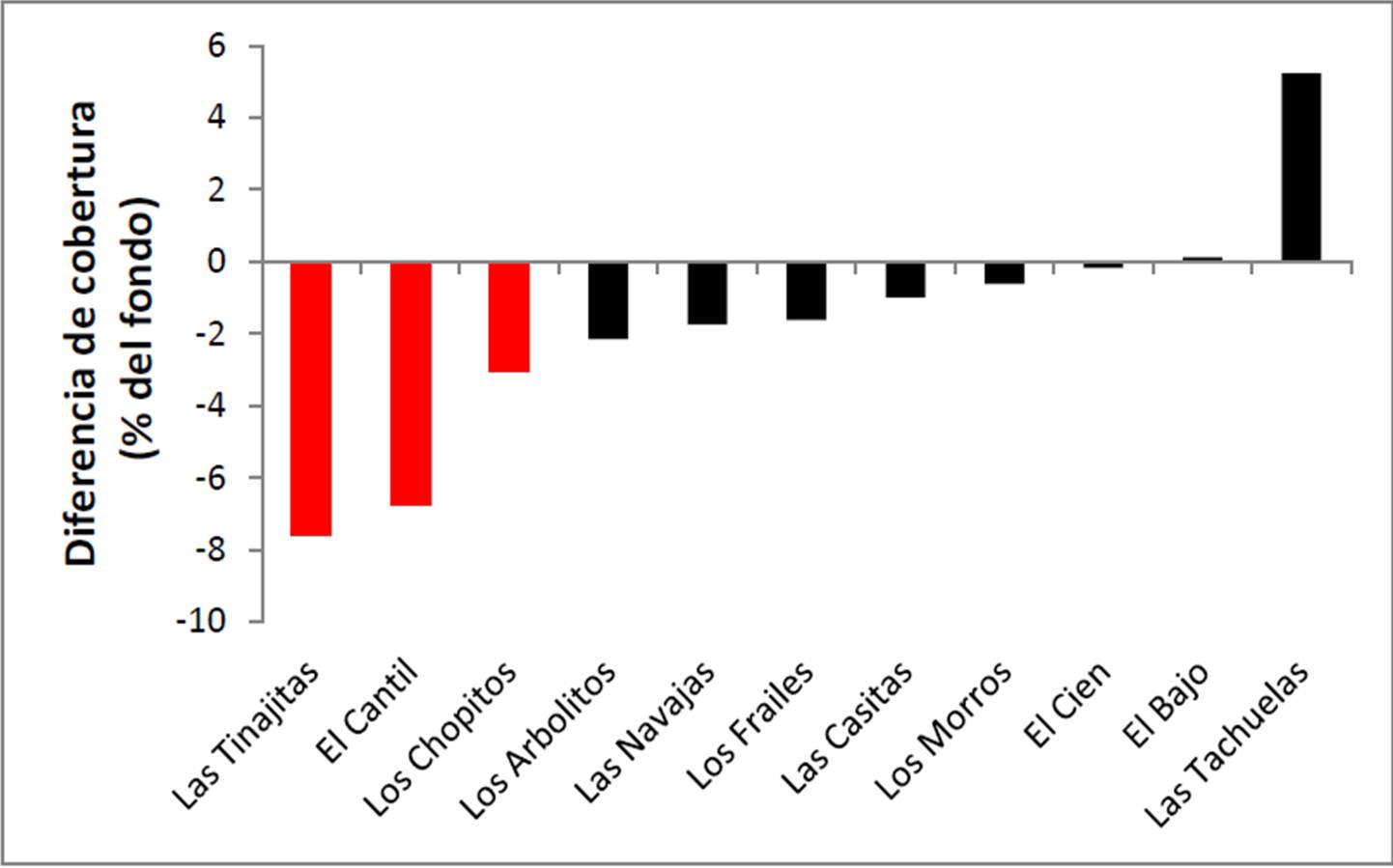 Figura 15. Cambio en la cantidad de cobertura viva en valores absolutos (porcentaje del fondo) entre agosto y noviembre de 2011 en el PNCP.
Figura 15. Cambio en la cantidad de cobertura viva en valores absolutos (porcentaje del fondo) entre agosto y noviembre de 2011 en el PNCP.
La literatura indica que el blanqueamiento de coral es normalmente causado por la elevación de la temperatura del mar a niveles que superan el rango de tolerancia de las zooxantelas, el cual se define como 1°C por encima de la temperatura promedio a largo plazo (normalmente 25 años) del mes más cálido. En el caso de Cabo Pulmo, ese nivel es de 30.3° C, un grado arriba de la media de agosto (29.3° C entre 1984 y 2010). Revisando la información sobre la temperatura observada en el PNCP durante 2011 puede verse que de finales de julio hasta octubre, el umbral de blanqueo fue excedido en la localidad (Figura 16).
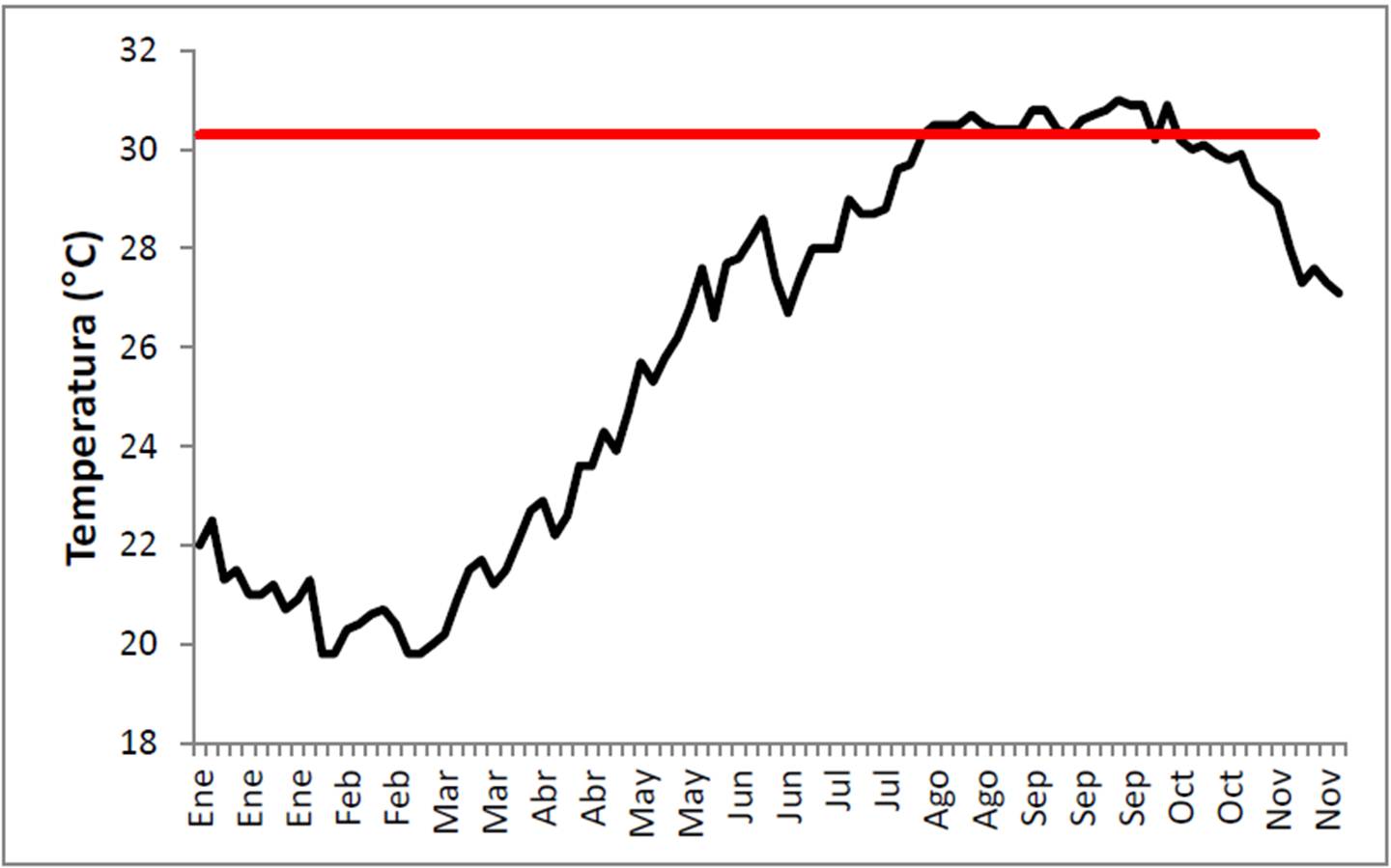 Figura 16. Temperatura superficial oceánica en el PNCP en los meses de Enero a Noviembre de 2011.
Figura 16. Temperatura superficial oceánica en el PNCP en los meses de Enero a Noviembre de 2011.
|
El que la temperatura marina rebase el umbral de blanqueo no causa daños inmediatos, sino acumulativos (Eakin et al. 2010). Se considera que si la elevación es de 1° C por semana o más, para la cuarta semana (28 días) de estar expuestos a estas condiciones los primeros corales van perdiendo color, para las 8 (56 días) semanas la pérdida de simbiontes es masiva y la muerte coralina inicia, y para las 12 semanas (84 días) la mortalidad es masiva e irreversible. En el PNCP, el día 1 de agosto señaló el inicio del stress térmico al alcanzarse los 30.3° C, y es por ello que para el momento de la visita inicial de este proyecto (16 a 19 de agosto), la decoloración solo era parcial. Para el 28 de agosto (temperatura de 30.4° C) deben haberse observado las primeras colonias completamente blanqueadas, mientras que para el 25 de septiembre (30.9° C) se habrían cumplido las 8 semanas de stress, marcando el inicio del blanqueo masivo. El día 13 de octubre por fin la temperatura llegó a 30.2° C, ya por debajo del umbral de blanqueo, sin embargo los corales ya habían estado poco más de 10 semanas (73 días en total) expuestos a temperaturas fuera de su límite de tolerancia. Para la fecha de la segunda visita al arrecife (13 al 16 de noviembre), la temperatura fluctuaba alrededor de 28° C y llevaba un mes de estar dentro de lo aceptable, pero el daño ocurrido debido a las condiciones de verano e inicios de otoño ya era patente, tal y como se cuantificó.
Además de lo anterior, hay que señalar que todo el año 2011 fue atípico, ya que sin que ocurrieran eventos de El Niño, a partir de mayo la temperatura fue significativamente más elevada que la normal en el sitio (Figura 17), y al menos tres meses estuvo arriba de los 30°C (agosto con 30.5°C, septiembre con 30.7°C y octubre con 30.2°C).
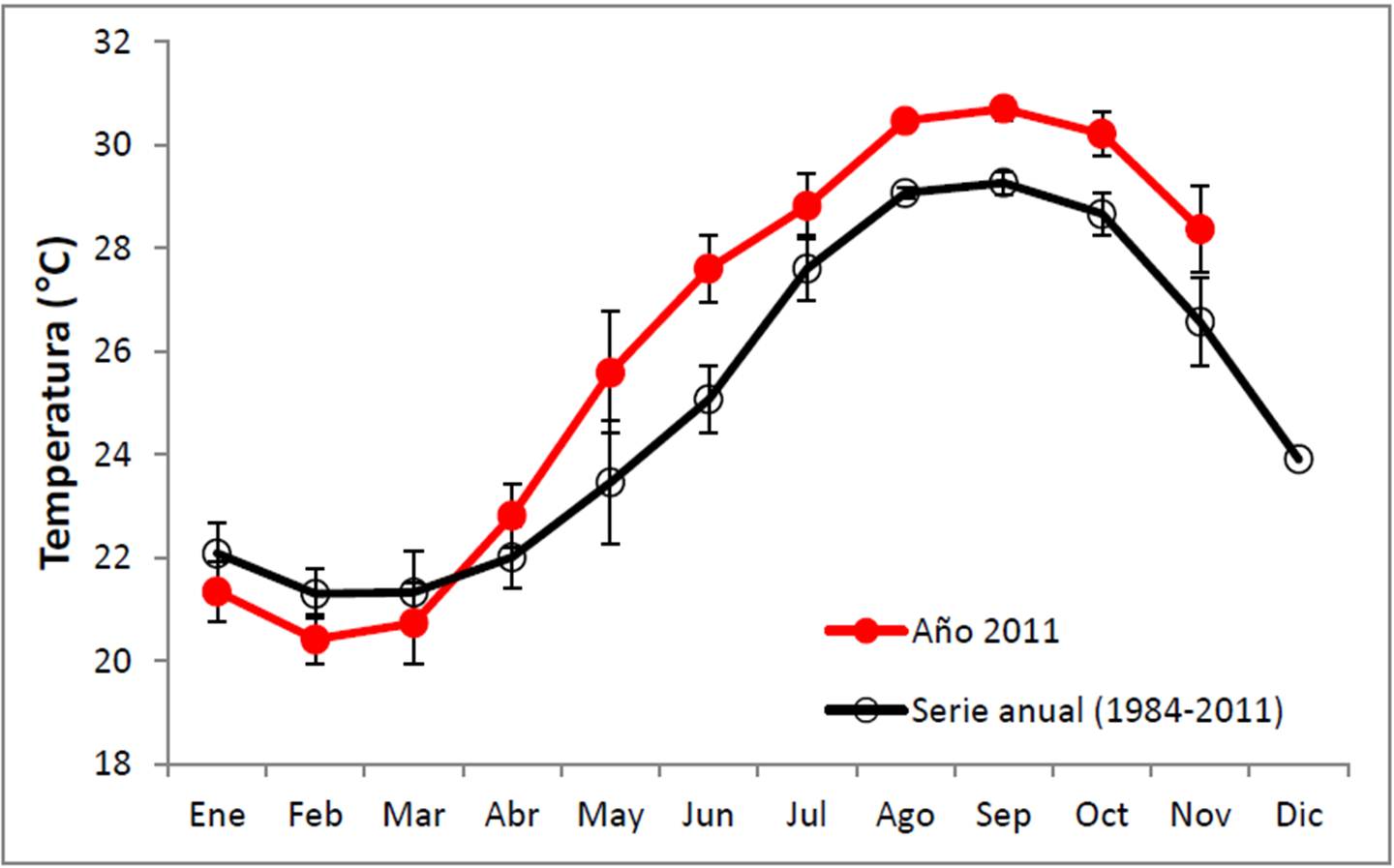 Figura 17. Comparación entre la temperatura superficial promedio mensual de 2010 en el PNCP, con la climatología local.
Figura 17. Comparación entre la temperatura superficial promedio mensual de 2010 en el PNCP, con la climatología local.
Las anomalías térmicas en 2011 fueron notables, tanto si se observan por día (Figura 18) o por mes (Figura 19). En el primer caso se ve que entre enero y marzo la temperatura estuvo ligeramente por debajo del promedio de 1984-2010 pero luego comenzó a elevarse hasta alcanzar un valor de +4.1° C el 30 de mayo. Anomalías superiores a +2° C siguieron presentándose casi todo junio (máximo de +3.5° C el 16 de junio), pero de julio a noviembre el valor fluctuó alrededor del +1.5° C, con un pico de +2.7° C el día 3 de noviembre. Finalmente, a partir del 17 de noviembre la temperatura regreso a la normalidad, con una anomalía registrada inferior a +1°C.
Como complemento a la información anterior, en la Figura 19 se denota que promediando por mes, junio y mayo presentaron los mayores valores de anomalías con +2.5° C y +2.1° C.
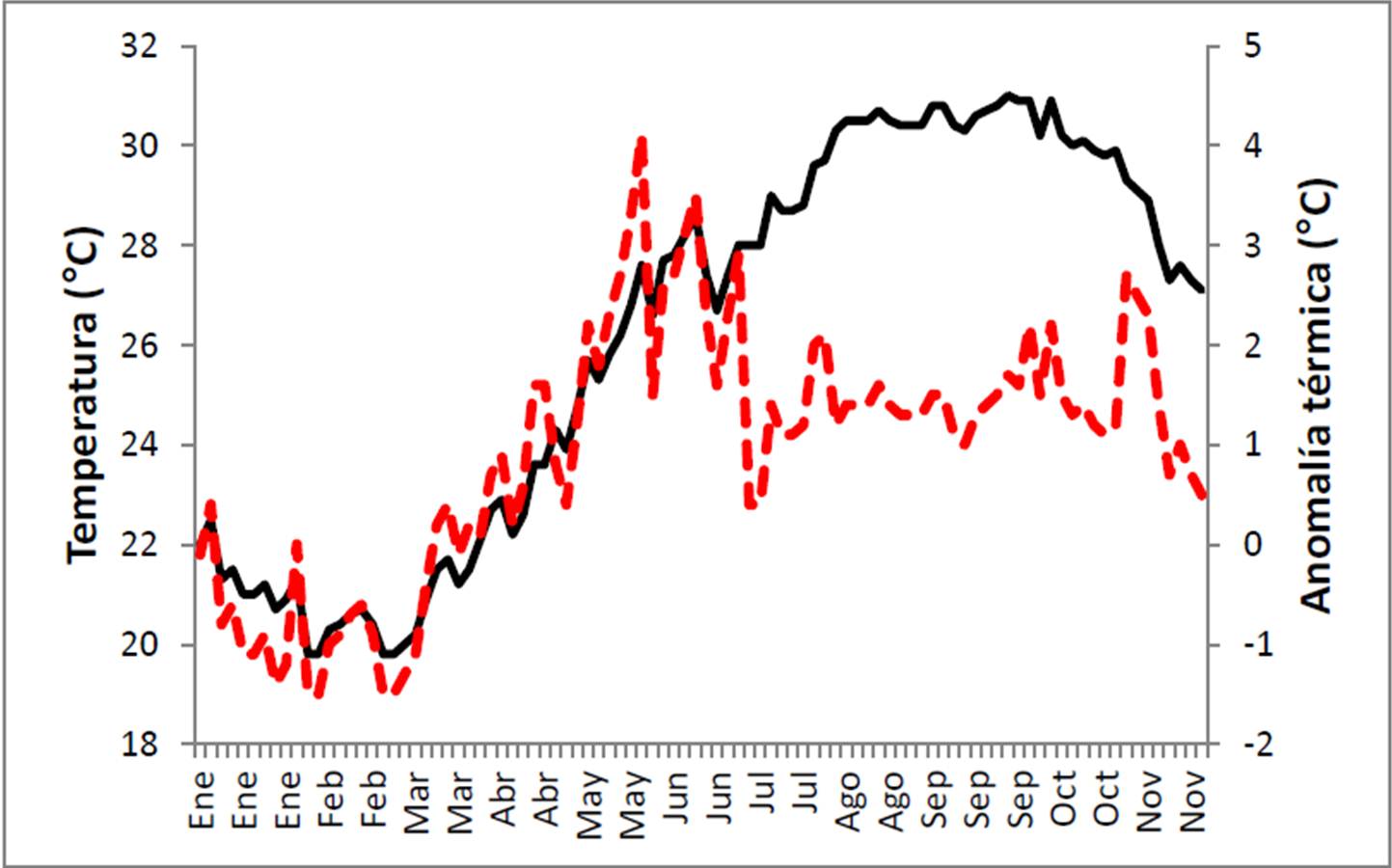 Figura 18. Anomalía térmica por día entre enero y noviembre de 2011 en el PNCP. Cálculos realizados con base en la climatología de 1984-2011 en el sitio.
Figura 18. Anomalía térmica por día entre enero y noviembre de 2011 en el PNCP. Cálculos realizados con base en la climatología de 1984-2011 en el sitio.
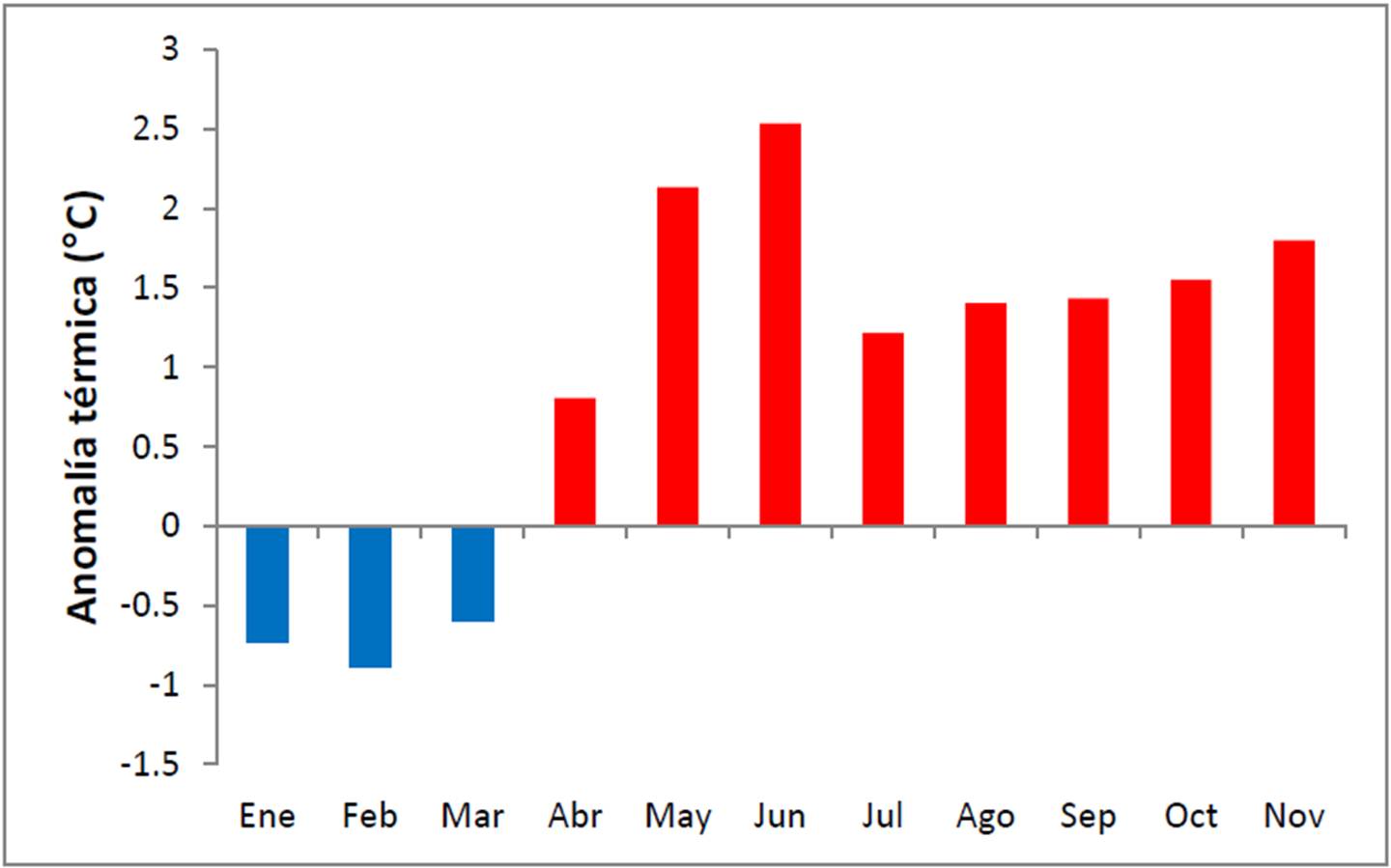 Figura 19. Anomalía térmica por mes entre enero y noviembre de 2011 en el PNCP. Cálculos realizados con base en la climatología de 1984-2011 en el sitio.
Figura 19. Anomalía térmica por mes entre enero y noviembre de 2011 en el PNCP. Cálculos realizados con base en la climatología de 1984-2011 en el sitio.
Para finalizar la sección, se pondrá el evento de blanqueamiento de 2011 en perspectiva con los ocurridos en la localidad en 1987 y 1997 (Figura 20). En el primer caso, Reyes Bonilla (1993b) menciona que se presentaron decoloraciones e incluso blanqueos totales, pero que la mortalidad no excedió el 10% de la población de Pocillopora spp., mientras que los demás géneros no fueron afectados. Ese año, el umbral de blanqueo se superó en solo septiembre con 30.4° C, aunque en agosto y octubre la temperatura superó los 29°C. La siguiente ocasión en que se alcanzó el umbral de pérdida de simbiontes fue en 1990 por un mes (septiembre; 30.3° C), pero el stress no causó daños mesurables. Sin embargo, en 1997, año en que se presentó el evento de Oscilación Sureña de El Niño más fuerte del Siglo XX (Reyes Bonilla et al. 2002), el límite de tolerancia se rebasó en agosto (30.8°C) y septiembre (30.4°C), mientras que en julio y octubre la temperatura superó los 29°C, y la anomalía en julio y agosto superó los 2°C. Para ese año, Reyes Bonilla (2001), menciona que en Cabo Pulmo hubo decoloraciones en julio, blanqueo masivo en octubre y regreso a las condiciones aceptables en noviembre. Sin embargo, lo más importante es que la mortalidad se presentó en los cuatro géneros de coral presentes en el arrecife, fue más alta en agua somera, y se perdió más del 50% de la cobertura viva, disminuyendo esta del 30% al 14% del fondo. Finalmente, el evento de 2011 descrito antes, causó mortalidades pero estas no pueden ser tomadas como elevadas, al ser inferiores al 3% de la cobertura total (equivalente al 14% de la población de corales en la localidad).
A manera de conclusión se afirma que el evento de blanqueo coralino de 2011 ha sido el segundo en relevancia en Cabo Pulmo desde 1984, pero no obstante ello, la pérdida de coral fue muy inferior a la observada en 1997. Esto es una clara evidencia de adaptación local a las condiciones de elevación de la temperatura resultado del cambio global, tal y como se demostró que ha ocurrido en diversos lugares del oeste de México (LaJeunesse et al. 2010), y en el Pacífico de Costa Rica (Guzmán y Cortés 2007).
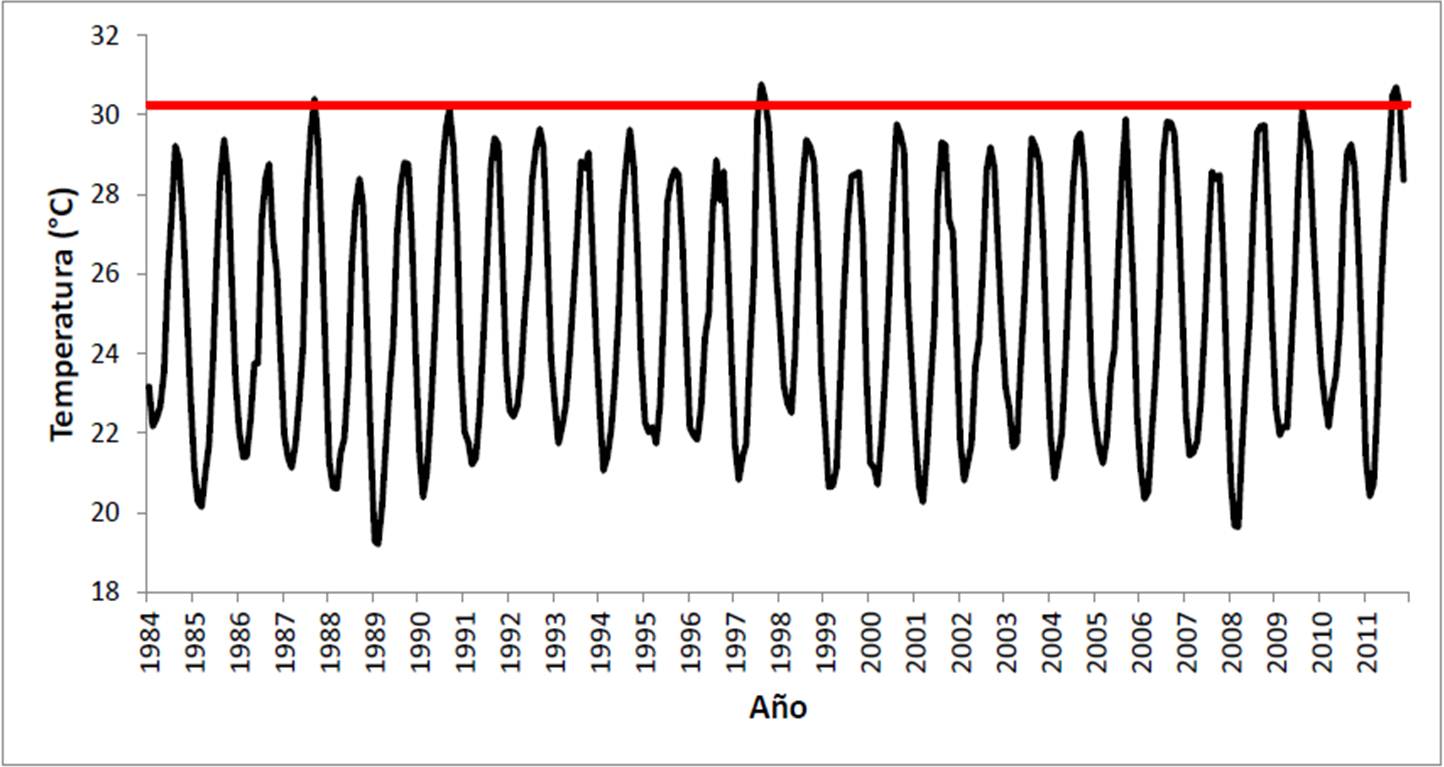 Figura 20. Temperatura mensual registrada en el PNCP entre 1984 y 2011. En rojo se marca el umbral de blanqueamiento de coral (30.2°C).
Figura 20. Temperatura mensual registrada en el PNCP entre 1984 y 2011. En rojo se marca el umbral de blanqueamiento de coral (30.2°C).
Para determinar el estado de conservación general de la estructura coralina del arrecife del PNCP, se tomó en cuenta todos los índices producidos y criterios de perturbación humana y natural utilizados en la presente investigación y se obtuvo un estimador del nivel de conservación de cada una de las áreas visitadas. Los Arbolitos, Las Navajas y El Cantil resultaron ser las tres áreas mejor conservadas de todo el parque, debido en especial al poco impacto humano aparente y baja mortalidad, y a su alta riqueza y diversidad de coral o por tener valores altos en los índices ecológicos, mientras que de manera contraria Los Frailes, Los Morros y Las Tachuelas figuran como las de peor estado por tener valores bajos en los índices ecológicos, alto blanqueo e impacto por fragmentación, el resto de las áreas se podrían considerar como en un nivel medio de conservación (Cuadro I y II).
Cuadro 1. Valores de los indicadores de salud coralina en el PNCP.
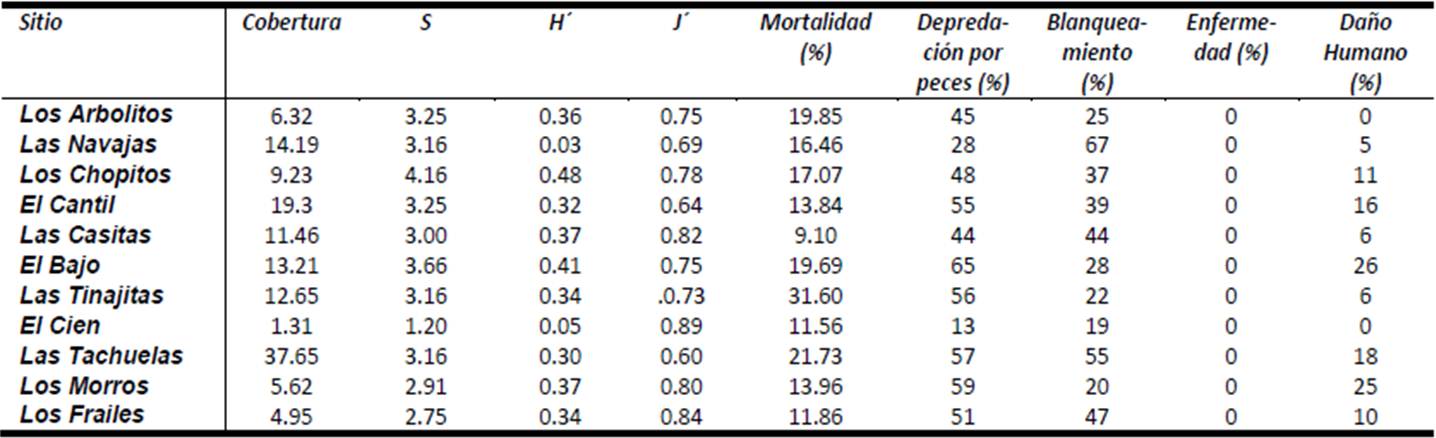
Cuadro 2. Valores ranqueados de los indicadores de salud coralina en el PNCP.
Para identificar las zonas coralinas susceptibles de restauración en el PNCP, como se observa en el Cuadro I, Los Frailes, Los Morros y Las Tachuelas, figuran como las tres áreas en peor estado de conservación, por lo que podrían ser consideradas buenas candidatas para su restauración. De ellas, Los Morros no presentan las mejores condiciones para considerarse una de las opciones a un programa de restauración coralina dada su profundidad (> 12 m), la cual no favorece al desarrollo de los corales. Algo similar podría decirse de El Cien, otro de los sitios con relativamente baja salud en el arrecife, bajo la óptica de los indicadores aquí empleados. Tomando lo anterior en consideración, Los Frailes, Las Tachuelas y Las Tinajitas podrían ser buenas áreas para llevar a cabo esfuerzos de recuperación por ser relativamente someras (< de 6 m) y la incidencia de luz es alta e ideal para el desarrollo coralino.
Es importante anotar aquí que las sugerencias previas no se hacen solo en función a la cobertura coralina actual, o al nivel de daño producido por el hombre o la alta temperatura, sino que la idea fue generar indicadores de vulnerabilidad (índices ecológicos) y presión (blanqueo, mortalidad, etc.) con el fin de tener un marco general de los sitios, y con base en él, buscar cuales una vez recuperados podrían ayudar en el futuro a mejorar la condición general del arrecife y sus funciones ecológicas.
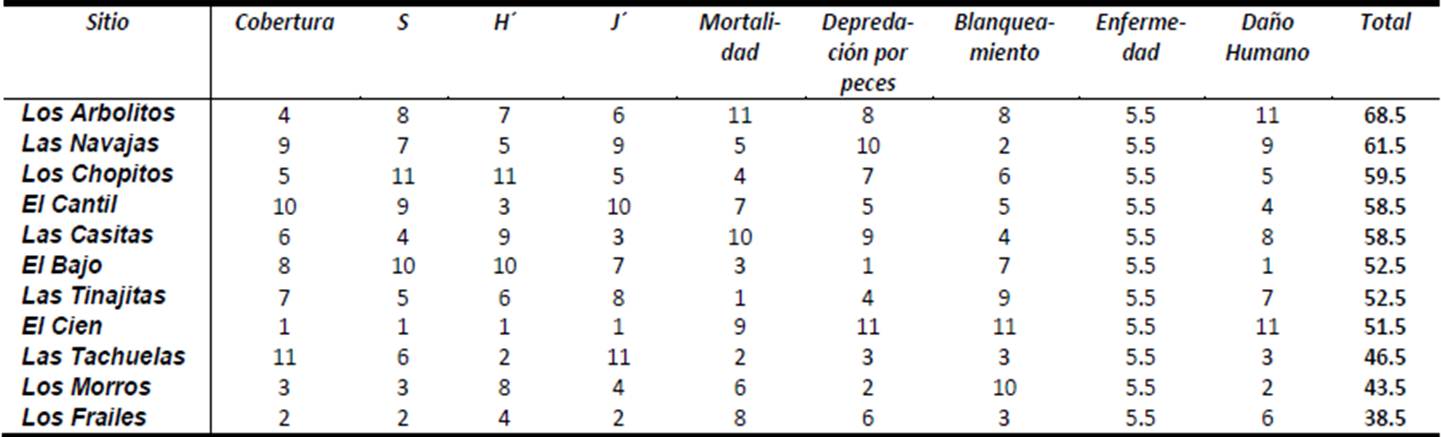
Según la información del satélite MODIS-AQUA y el SEAWIFS, el Parque Nacional Cabo Pulmo presenta promedios anuales de clorofila de 0.461 ± 0.098 mg/m3, encontrándose los valores más bajos para los años 2003 y 2008, mientras que 2007 tuvo el valor promedio más alto en los últimos doce años con 0.647 mg/m3 (Figura 21). El coeficiente de extinción de la luz (promedio general de 0.066 ± 0.007 1/m; Figura 22), sigue el mismo patrón de la clorofila, exhibiendo en general valores elevados en 2007 y 2000-2001, y bajos en 2003. Por otra parte, la profundidad de incidencia luminosa del 1% (indicador de la máxima profundidad de vida para un coral zooxantelado; Figura 23) mantiene un promedio de 16.61 ± 1.51 m, apreciándose que en 2003 hubo la mayor penetración de luz (19 m de promedio anual). Para 2007, por el contrario, se observa el valor promedio más bajo de incidencia (14 m). Finalmente, la Figura 24 muestra que la temperatura ha sido notablemente estable de 2003 a 2010 (promedio de 25.02 ± 0.95 °C) aunque en 2002 fue más cálida.
A manera de resumen, al menos entre 1998 y 2010 no parece haber cambios notables en la producción primaria superficial, ni una elevación tangible de la temperatura del PNCP; quizá en parte debido al hecho de que para el Pacifico Oriental, las condiciones se han hecho ligeramente más frías a partir del inicio de la década, producto de la Oscilación Decadal del Pacifico en tránsito, hacia su fase fría (Lluch Cota et al. 2010).
Sin embargo, donde se observan notables modificaciones interanuales es en el coeficiente de extinción de luz y por ende en la penetración luminosa hacia la profundidad. Si bien ambas fluctúan alrededor de sus valores promedio de 0.066 ± 0.007 1/m y 16.61 ± 1.51 m, respectivamente, hay años como 2000, 2001 y 2007 cuando la incidencia solo llega a -15 m incluso menos; esto implica que los corales de zonas como El Bajo o Los Morros pueden ver afectada su actividad metabólica por falta de energía para sus simbiontes. Eventos de este tipo pueden disminuir sensiblemente la tasa de crecimiento coralino, así como el esfuerzo reproductivo de las colonias (Harrison 2010). Si bien lo mencionado es un fenómeno natural, es importante poner en perspectiva el hecho que cambios en el uso de suelo derivan en aumentos de turbidez (descarga o agitación de sedimentos, cambios en la salinidad) y que elevaciones en la refractancia luminosa o el aumento de la productividad primaria en la columna de agua debidos a la presencia de altos nutrientes, pueden tener un efecto considerable sobre el ecosistema.
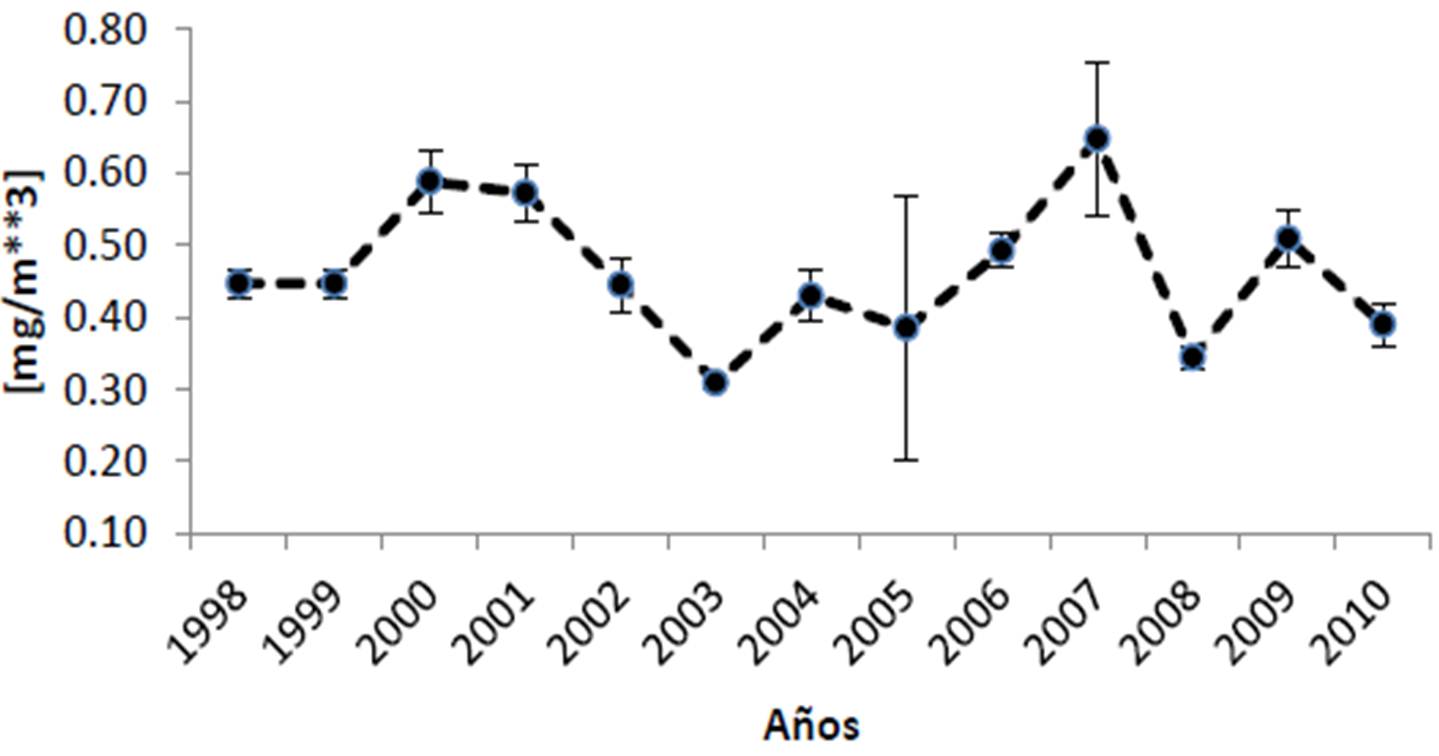
Figura 21. Promedio anual de concentración de clorofila de 1998 a 2010 en el PNCP.
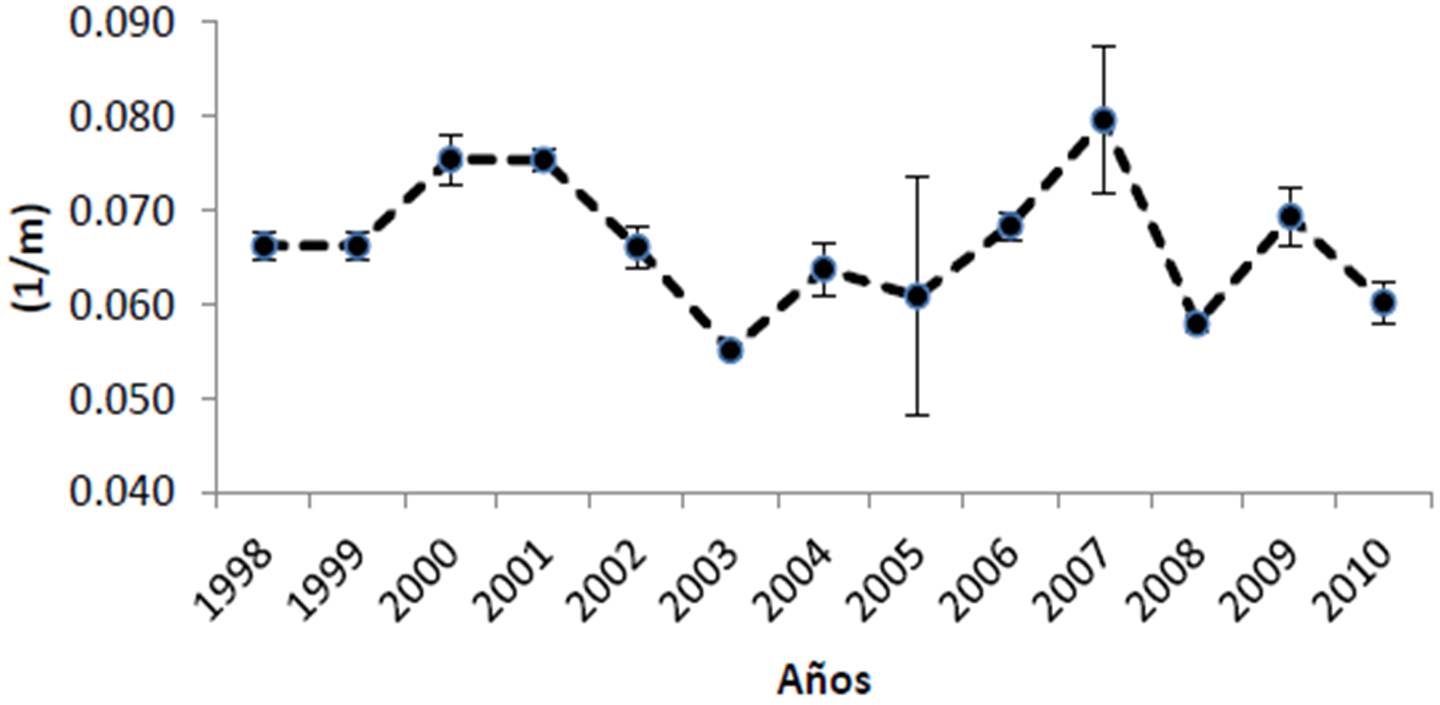 Figura 22
Figura 22. Promedio anual del coeficiente de extinción de la luz de 1998 a 2010 en el PNCP.
 Figura 23
Figura 23. Promedio anual de la profundidad a la que se presenta el 1% de incidencia luminosa de 1998 a 2010 en el PNCP.
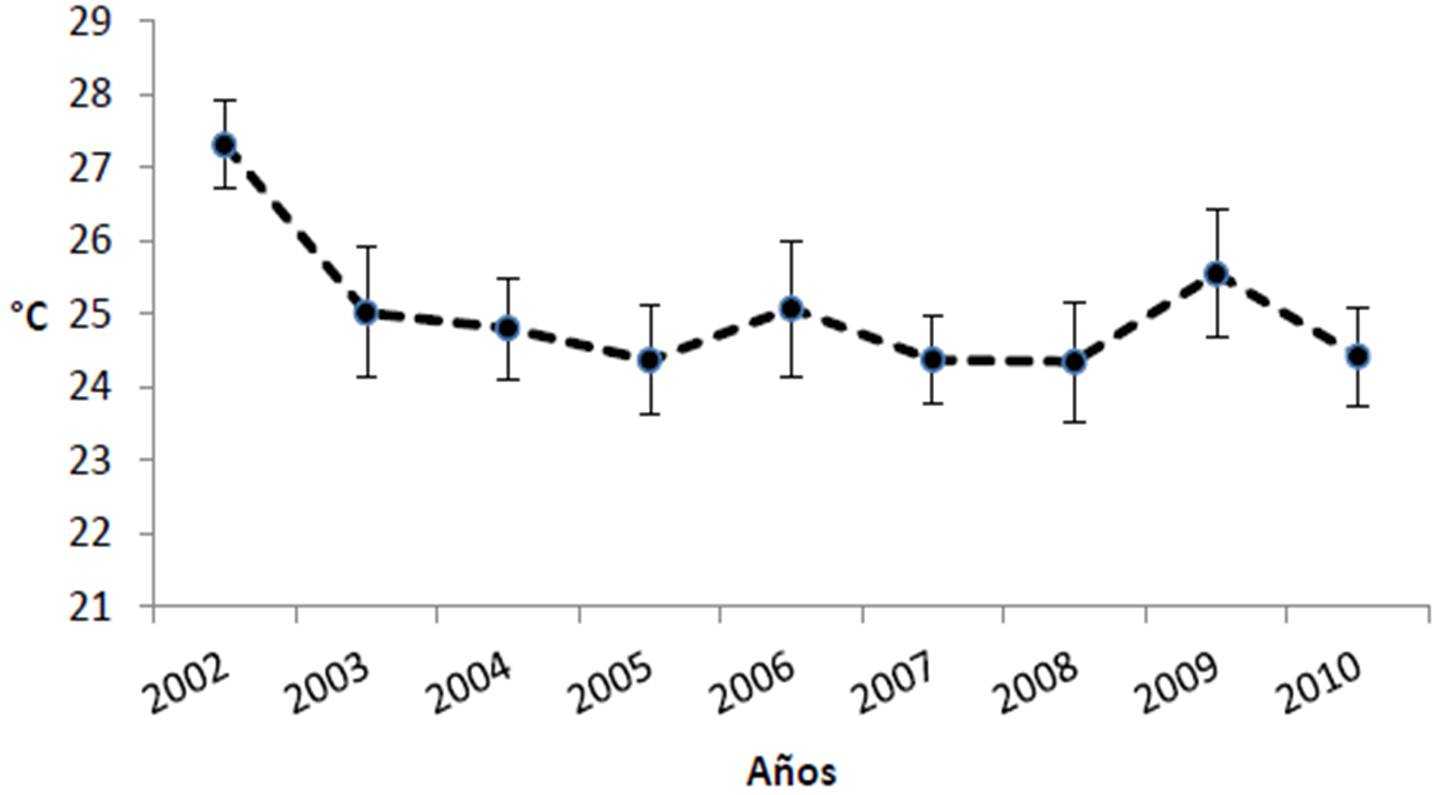
Figura 24. Promedio anual de temperatura superficial de 1998 a 2010 en el PNCP.
Analizando ahora por meses del año, el promedio mensual de clorofila en el Parque Nacional Cabo Pulmo es de 0.459 ± 0.064 mg/m3, siendo enero y diciembre los meses que muestran los valores más altos con 0.69 y 0.67 mg/m3 respectivamente, y agosto y octubre los más bajos (0.23 y 0.21 mg/m3; Figura 25). Es notable como cada año entre noviembre y diciembre la cantidad de clorofila se duplica en promedio. Mismo patrón temporal se presenta para el coeficiente de extinción de la luz (Figura 26) con un promedio de 0.066 ± 0.004 1/m, la incidencia luminosa (promedio general de 16.73 ± 0.99 metros; Figura 27) es superior en octubre con un alcance de 23 m, la menor penetración de luz en el PNCP se presentó en enero y diciembre con 12.1 y 12.6 metros respectivamente. Es interesante que como resultado de la disminución abrupta de la transparencia del agua de noviembre a diciembre, la profundidad de penetración de la luz disminuye 7 metros en promedio. En cuanto a la temperatura (Figura 28), se presenta un promedio anual de 24.73 ± 3.17 °C, siendo agosto (28.6°C), septiembre (28.9°C) y octubre (28.7°C) los más cálidos, febrero y marzo son los más fríos (20.7°C).
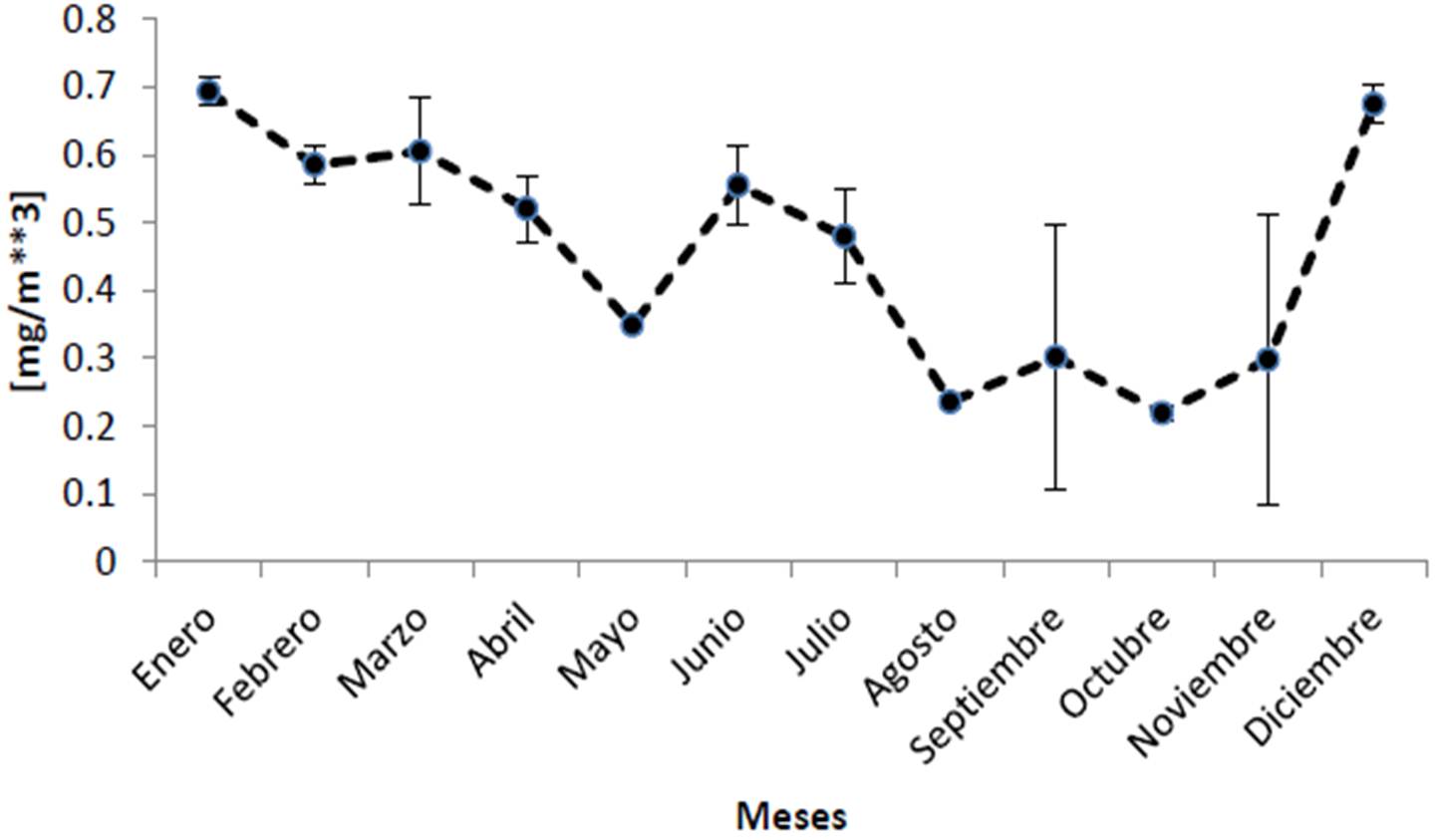 Figura 25. Concentración mensual (promedio ± error estándar) de clorofila en el PNCP.
Figura 25. Concentración mensual (promedio ± error estándar) de clorofila en el PNCP.
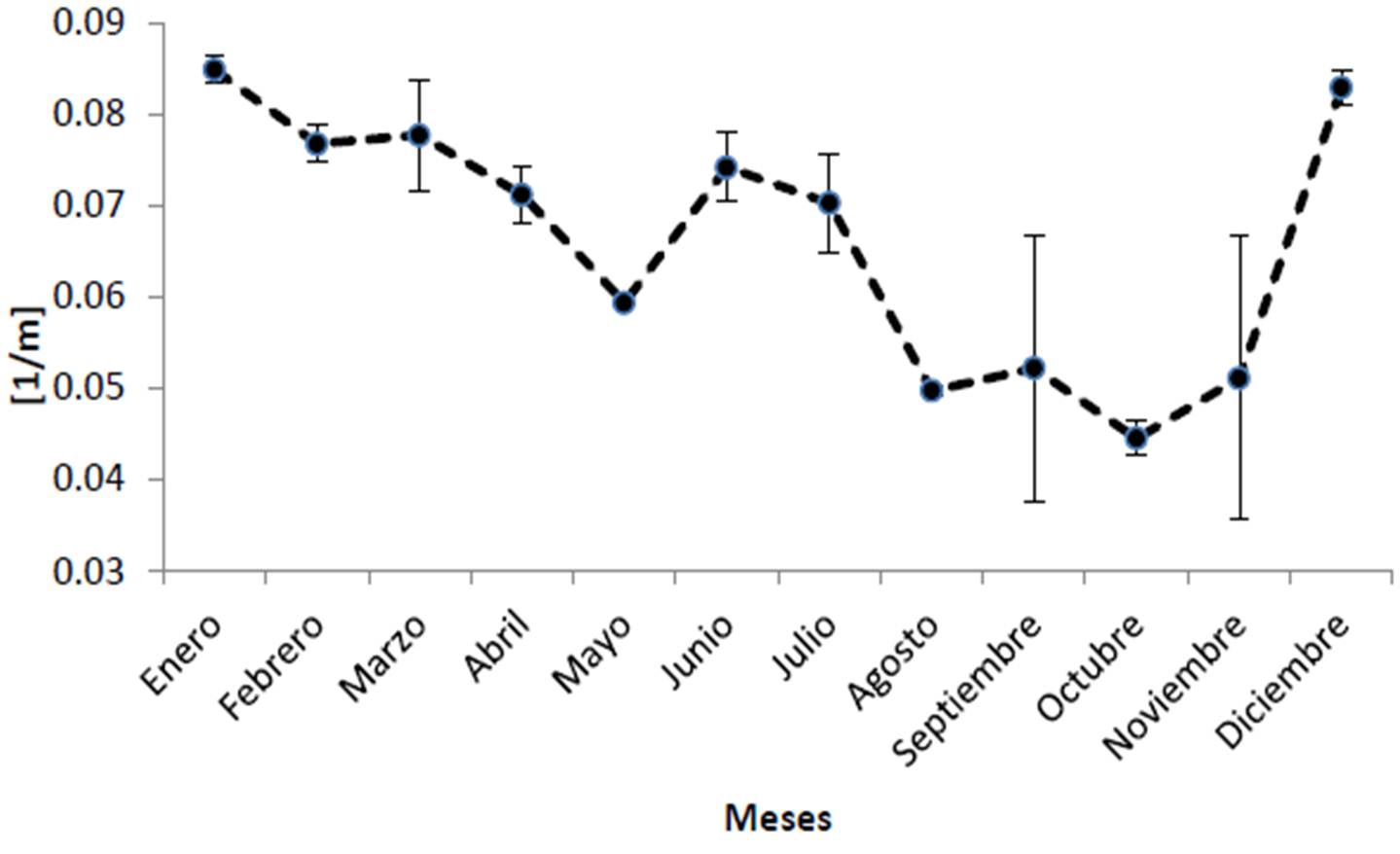 Figura 26. Concentración mensual (promedio ± error estándar) de clorofila en el PNCP.
Figura 26. Concentración mensual (promedio ± error estándar) de clorofila en el PNCP.
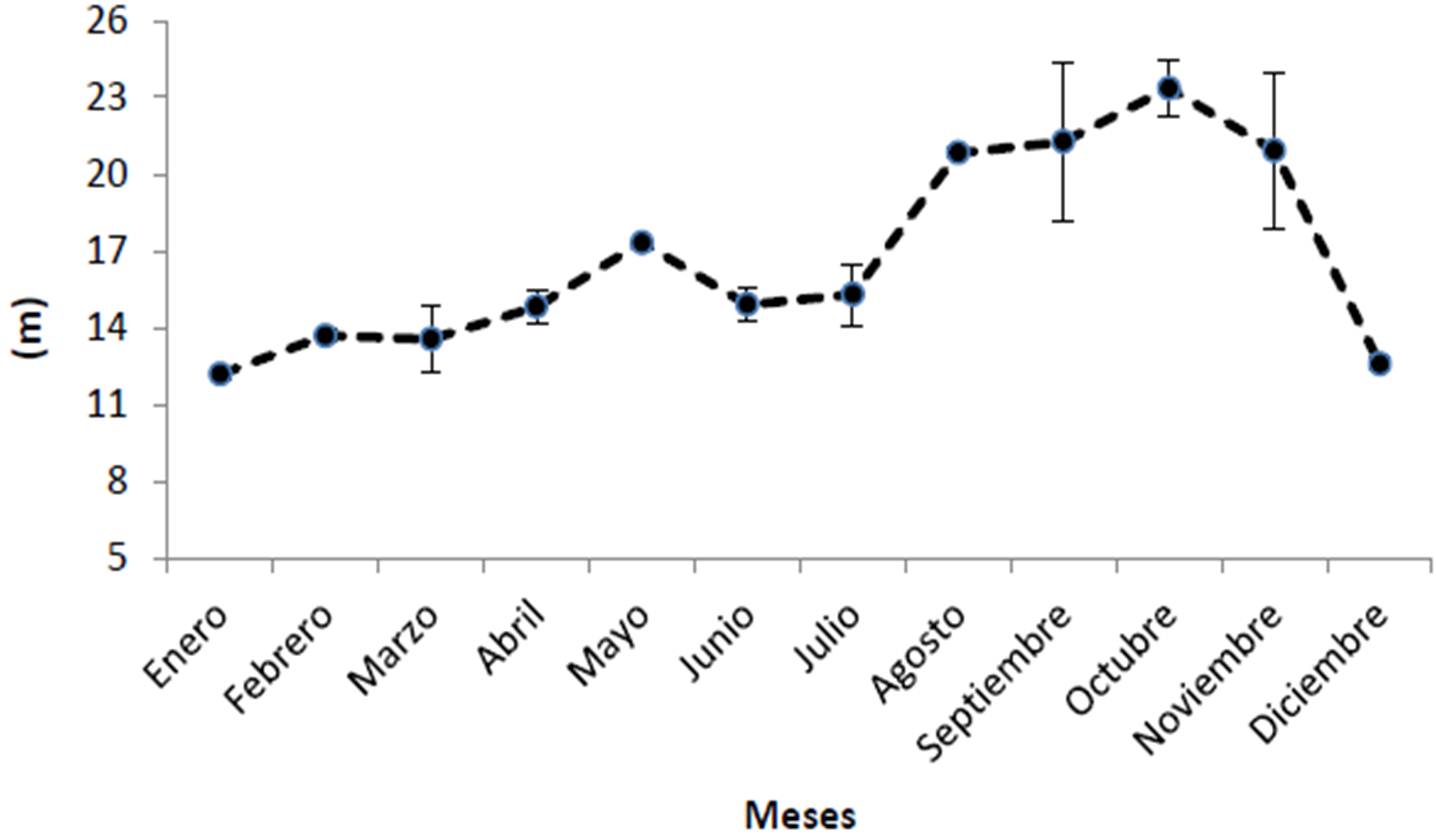 Figura 27. Promedio mensual de la profundidad a la que se presenta el 1% de incidencia luminosa en el PNCP.
Figura 27. Promedio mensual de la profundidad a la que se presenta el 1% de incidencia luminosa en el PNCP.
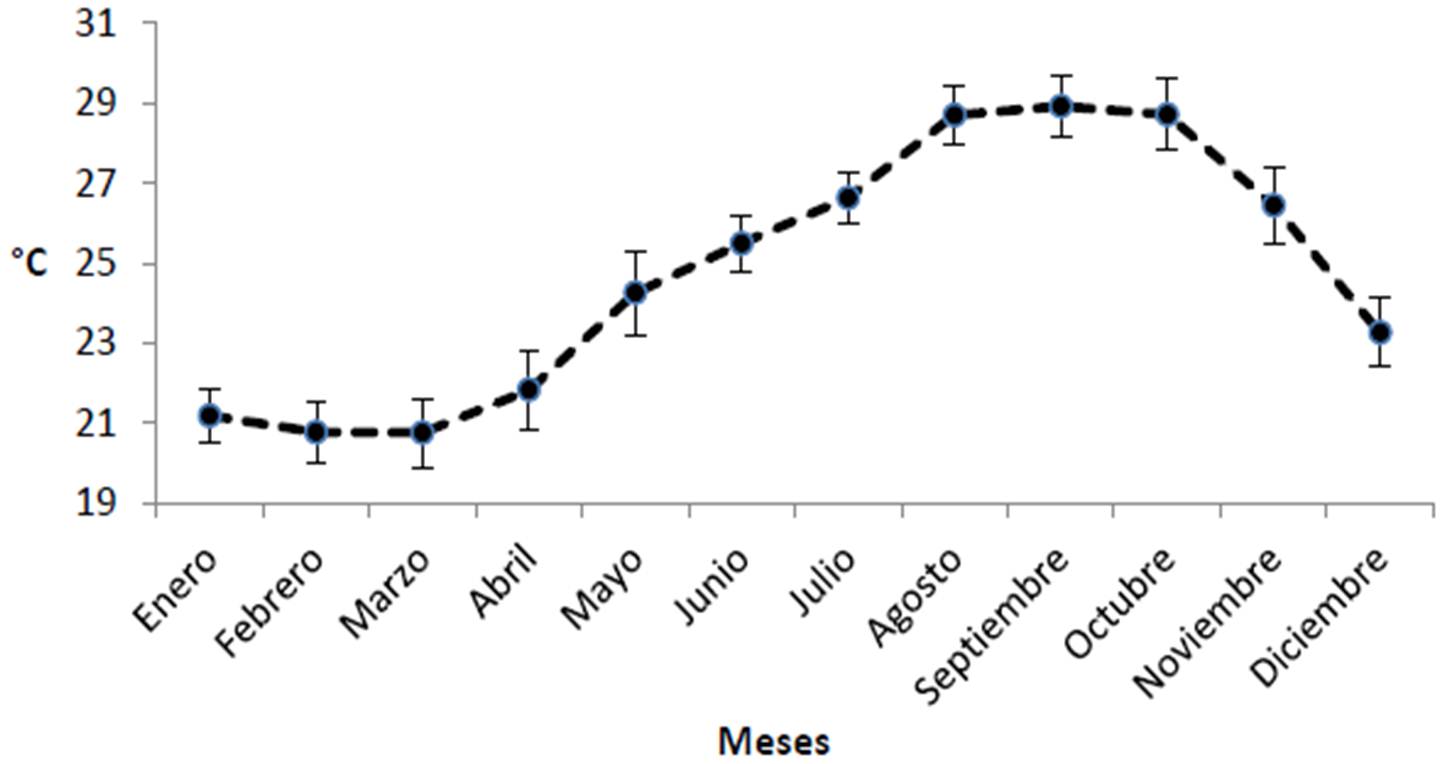 Figura 28. Promedio mensual de temperatura en el PNCP.
Figura 28. Promedio mensual de temperatura en el PNCP.
El patrón general a escala espacial de las condiciones ambientales del PNCP se muestra en la Figura 29 (a, b y c), en donde las concentraciones de clorofila van desde los 0.40 mg/m3 hasta los 0.58 mg/m3, presentándose los valores más bajos al sureste, y los más altos al suroeste de la región, es decir alrededor de Los Frailes. En este caso el resultado se debe al efecto de elevación local de nutrientes que resulta del movimiento de agua a lo largo del Cañón de Los Frailes, el cual origina una surgencia continua en la porción sur del parque. Por su parte, la profundidad del 1% de luz presenta un patrón opuesto pues Los Frailes y la porción sur de la bahía tienen aguas menos transparentes que las del océano abierto o en el extremo norte del PNCP. Finalmente y en cuanto a la temperatura, se observa una clara tendencia de aumento hacia el este de la región, con un intervalo de 5°C aproximadamente desde la zona menos cálida (sur-oeste) hasta la más tórrida (sur-este). También se denota en este factor el efecto de enfriamiento del Cañón de Los Frailes.
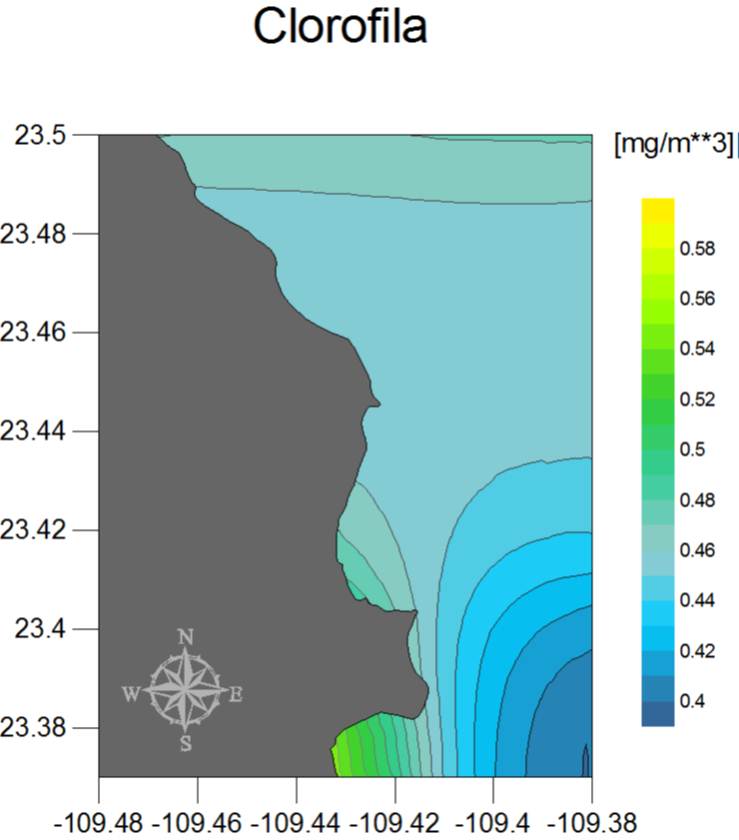 Figura 28a.
Figura 28a.
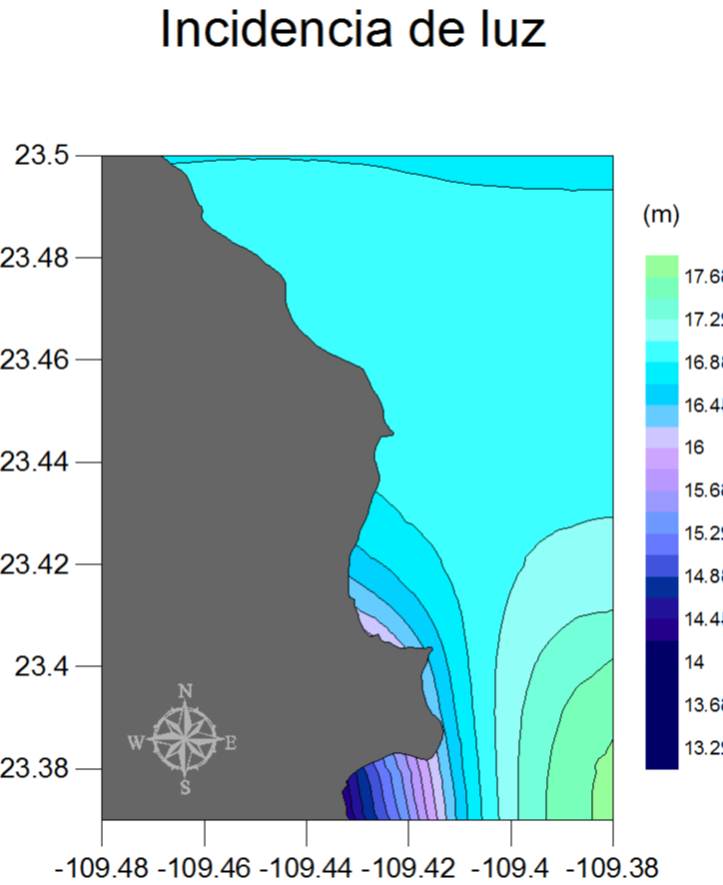 Figura 28b
Figura 28b.
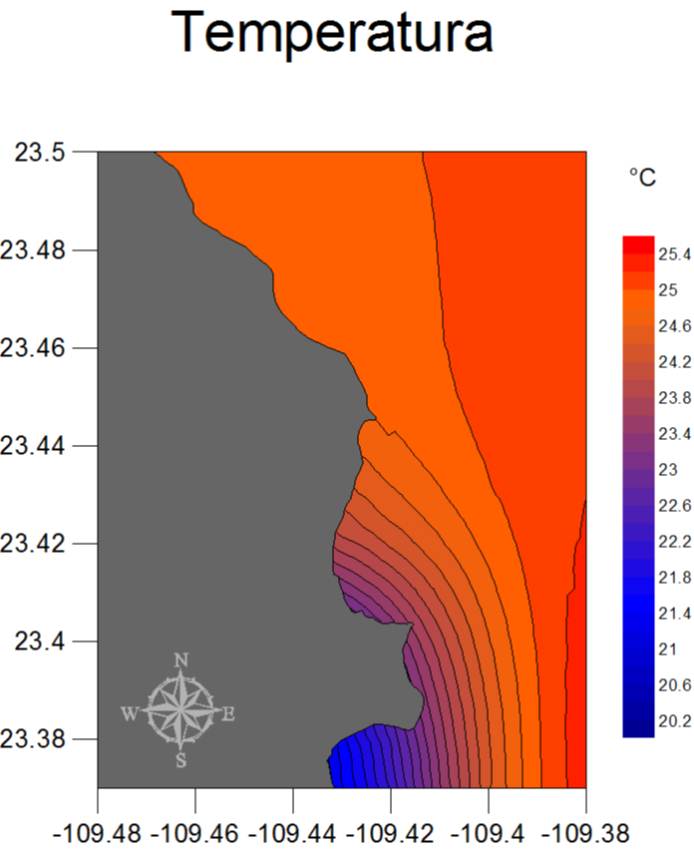
Figura 28c. Mapas de promedios anuales de las condiciones ambientales en el PNCP (de 1998 a 2010 para clorofila e incidencia de luz, y de 2002 a 2010 para la temperatura).
Discusión y Conclusiones: Se encontraron 9 especies de coral en el PNCP, pertenecientes a los géneros Pocillopora (5 especies), Pavona (2 especies), Porites y Psammocora (1 especie), con un promedio de 3.9 spp/censo. La zona más rica del parque fue Los Chopitos (4.1 spp/censo).
La cobertura coralina en el PNCP es de 12.46%, siendo más alta en Las Tachuelas (37.7%) y El Cantil (19.3%). Si solo se consideran sitios relativamente someros donde los corales pueden habitar sin problemas de baja luminosidad (<-10 m), la cobertura pasa a ser del 14.6% del fondo. Las especies dominantes fueron Pocillopora meandrina y P. verrucosa, con un 34% y 33% de la cobertura total, respectivamente.
La diversidad coralina promedio fue de 0.34 decits, y los sitios más diversos fueron Los Chopitos (0.34 decits), seguido por El Bajo. La equidad promedio fue de 0.74 unidades, y la zona con mayor valor de este índice fue El Cien (0.89 unidades).
En 2010 el porcentaje de fondo arenoso era del 56%, de coral vivo 21%, de coral muerto 11% y de pavimento rocoso 6%, siendo estos los cuatro tipos predominantes de hábitat bénticos en el PNCP.
La incidencia de enfermedades en el parque fue de cero, la fragmentación aparentemente de origen humano fue del 12% de las colonias (más alto en El Bajo, Los Morros y Las Tachuelas, sitios de frecuentes visitas por buceadores), y el daño por depredación de peces fue del 50%, especialmente en zonas al norte del parque como El Bajo (64% de corales mordidos), Los Morros (58%) y Las Tachuelas (57%).
Entre agosto y noviembre de 2011 se presentó un evento de blanqueamiento importante en la zona de estudio. La mortalidad final fue del 2.4% de la cobertura coralina sobre el fondo (equivalente al 14% de la población total). Esta cifra es notablemente menor que la registrada para el blanqueamiento de 1997, cuando se perdió más del 50% de la cobertura, con una elevación de temperatura similar. La diferencia citada apunta hacia la exitosa adaptación local de las comunidades coralinas de Cabo Pulmo a las altas temperaturas, como ha ocurrido a lo largo del Pacífico oriental en años recientes.
La causa del blanqueamiento fue la elevada temperatura del mar, que sobrepasó el umbral de pérdida de zooxantelas (30.3°C) entre agosto y octubre (73 días). Las zonas más afectadas fueron Las Tachuelas (71% de las colonias), Las Navajas (66%) y Los Frailes (61%). Como resultado del evento la mayor pérdida de abundancia de coral ocurrió en Las Tinajitas (-7.6%), El Cantil (-6.7%) y Los Chopitos (-3.1%).
Tomando en cuenta 8 indicadores de la salud coralina (riqueza, abundancia, diversidad, equidad, porcentaje de mortalidad colonial, depredación por peces, incidencia de blanqueamiento, enfermedades y daños por acción humana), las zonas mejor conservadas en el PNCP fueron Los Arbolitos, Las Navajas y El Cantil, mientras que Los Frailes, Los Morros y Las Tachuelas presentaron el estado menos apropiado.
Tomando en cuenta la profundidad de los sitios y su condición actual, se recomienda que un programa de restauración coralina se inicie en las zonas de Los Frailes, Las Tachuelas y Las Tinajitas.
Los valores promedio de los parámetros oceanográficos analizados fue la siguiente: la concentración de clorofila a en la columna de agua de 0.647 mg/m3, el coeficiente de extinción de la luz de 0.066, la profundidad de 1% de luz de 16.6 m, y la temperatura de 25.02°C.
A escala mensual, en invierno y primavera (diciembre a junio) se alcanzan los más altos valores de clorofila a y del coeficiente de extinción de la luz, mientras que entre agosto y noviembre ocurren los más bajos. Situación inversa, la profundidad del 1% de luz y temperatura son más elevadas en verano-otoño, que el resto del año.
Finalmente, la clorofila y la transparencia del agua no son homogéneas en el arrecife, sino que son más altas en la zona circundante a Los Frailes por efecto de la surgencia local causada por el cañón. En contraste, la temperatura media es más baja en esa zona que en el resto del área de estudio.
Recomendaciones: El seguimiento a las condiciones de la comunidad coralina del Parque Nacional Cabo Pulmo es clave para apoyar las labores de manejo y toma de decisiones del área protegida, además cobra relevancia debido al evento de blanqueamiento que se presentó en 2011 y que afectó el 14% de la población coralina total en el arrecife. En 1997, cuando también se presentó un fenómeno semejante, el monitoreo de la situación no fue llevado a cabo dada la carencia de autoridades designadas en el parque; con ello se perdió una gran oportunidad para evaluar el efecto relativo del evento tanto en la comunidad coralina como en el ecosistema en su conjunto. Afortunadamente en 2011 ya se cuenta con un staff para la administración del sitio, y fondos para realizar las labores que permitan evaluar la situación arrecifal en el futuro.
Con base en lo anterior y a la experiencia obtenida durante el proyecto PROMOBI “Monitoreo de corales en el Parque Nacional Cabo Pulmo, B.C.S”, se hacen las siguientes sugerencias para mejorar la calidad de la información a generarse en futuras campañas de monitoreo en el parque:
- Realizar el monitoreo en campo que cubra las dos épocas del año (temporada fría entre diciembre y mayo, y temporada cálida de junio a noviembre), en lo posible llevando a cabo la misma metodología y conduciendo al menos 5 censos por sitio/temporada, o al menos 50 en total en el parque por temporada.
- Mantener el seguimiento en los sitios actualmente elegidos para los censos, pero incluir otras zonas de interés como podrían ser el Pecio “El Vencedor”, El Islote, Cabo Pulmo, La Esperanza y los parches coralinos entre Las Tachuelas y la entrada de la bahía.
- Si lo anterior no es posible, seleccionar los sitios de monitoreo con base en un sistema estratificado que incluya tanto zonas someras como profundas, y que realice un número de censos proporcional a la cobertura de coral por sitio.
- Tomar datos sobre los mismos parámetros biológicos que se consideraron en 2011 (cobertura de coral, incidencia de blanqueamiento, enfermedades, daño por el hombre, etc.), ya que permiten tener una idea clara de la situación de las comunidades bénticas en el arrecife. Complementar esta información con registros de heterogeneidad del fondo y de abundancia de invertebrados asociados al bentos (equinodermos, moluscos, peces crípticos). Estos datos permitirán mejorar la precisión de los análisis comunitarios, y las estimaciones del nivel de conservación de los sitios.
- Compaginar los sitios de monitoreo coralino con los de monitoreo ictiológico llevado a cabo por Comunidad y Biodiversidad A. C. para tener indicadores más robustos y menos parciales de la condición real del PNCP.
- En el caso que residentes locales apoyen en la labor de monitoreo, tratar que al menos la mitad de los censadores tengan un trasfondo académico. Esto podría asegurar el nivel de calidad de la información, el compromiso con la labor, y ayudará a reducir el tiempo de captura de los datos, de modo que las autoridades puedan tener disponible la información a corto plazo.
- Colocar una serie de medidores de temperatura y luz (“hobos”) en al menos seis sitios del PNCP, siendo dos dentro de la bahía, dos al norte y dos al sur. La idea es que estos sensores ofrezcan información sobre el perfil batimétrico de los parámetros oceanográficos.
- Durante los patrullajes semanales en el parque, tomar registros de temperatura superficial y profundidad del disco de Secchi en algunos de los sitios de monitoreo coralino. Esto servirá para realizar correcciones a los datos satelitales de las condiciones térmicas y de transparencia del agua (coeficiente de extinción de la luz y profundidad del 1% de intensidad luminosa).
- Mantener anualmente la búsqueda de información oceanográfica por satélite para actualizar la base de datos entregada a las autoridades del parque.
|
 Buzón de sugerencias y comentarios
Buzón de sugerencias y comentarios